අපි හැමෝටම කුඩා කාලයේ ඉගැන්වූයේ ඉන්ද්රිය පහක් ඇති බවයි: පෙනීම, රසය, ශබ්දය, සුවඳ සහ ස්පර්ශය. ආරම්භක ඉන්ද්රියයන් හතර ඇස්, රස අංකුර, කන් සහ නාසය වැනි පැහැදිලි, වෙනස් ඉන්ද්රියයන් භාවිතා කරයි, නමුත් ශරීරය සංවේදනය හරියටම ස්පර්ශ කරන්නේ කෙසේද? ස්පර්ශය අභ්යන්තරයේ මෙන්ම පිටතද මුළු ශරීරයම අත්විඳියි. ස්පර්ශ සංවේදනය සඳහා වගකිව යුතු එක් විශේෂිත ඉන්ද්රියයක් නොමැත. ඒ වෙනුවට, මුළු සිරුර වටා කුඩා ප්රතිග්රාහක හෝ ස්නායු අවසානයක් ඇති අතර එමඟින් ස්පර්ශය සිදුවන ස්ථානය දැනෙන අතර එය සිදු වූ ස්පර්ශයේ වර්ගය පිළිබඳ තොරතුරු මොළයට සංඥා යවයි. දිවේ ඇති රස අංකුරයක් රසය හඳුනා ගන්නා පරිදි, යාන්ත්රික ප්රතිග්රාහක යනු සමේ සහ අනෙකුත් අවයවවල ස්පර්ශ සංවේදනයන් හඳුනා ගන්නා ග්රන්ථි වේ. ඔවුන් ලෙස හැඳින්වේ යාන්ත්රික ප්රතිග්රාහක මක්නිසාද යත් ඒවා නිර්මාණය කර ඇත්තේ යාන්ත්රික සංවේදනයන් හෝ පීඩනයේ වෙනස්කම් හඳුනා ගැනීමට ය.
යාන්ත්රික ප්රතිග්රාහක භූමිකාව
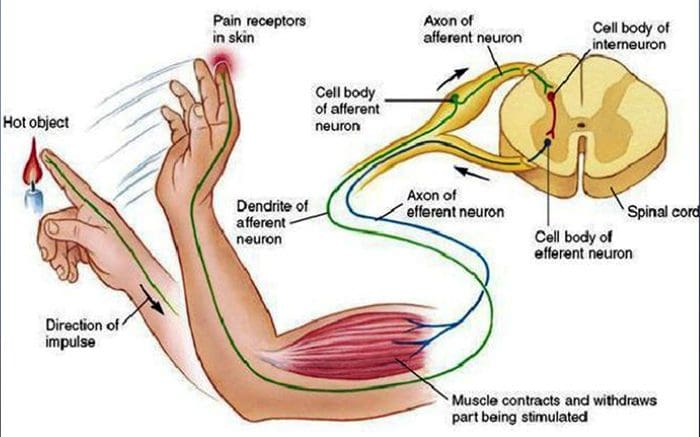
එම නිශ්චිත ඉන්ද්රිය සොයා ගැනීමට වගකිව යුතු ඉන්ද්රිය මොළයට පණිවිඩයක් යවන විට තමන් සංවේදනයක් අත්විඳ ඇති බව පුද්ගලයෙකු තේරුම් ගනී, එය සියලු තොරතුරු සැකසීමට සහ සකස් කරන මූලික ඉන්ද්රිය වේ. නියුරෝන ලෙස හඳුන්වන වයර් හරහා ශරීරයේ සියලුම ප්රදේශවලින් මොළයට පණිවිඩ යවනු ලැබේ. මිනිස් සිරුරේ සෑම ප්රදේශයකටම විහිදෙන කුඩා නියුරෝන දහස් ගණනක් ඇති අතර, මෙම නියුරෝන බොහොමයක අවසානය මත යාන්ත්රික ප්රතිග්රාහක වේ. ඔබ වස්තුවක් ස්පර්ශ කරන විට සිදු වන දේ නිරූපණය කිරීම සඳහා, අපි උදාහරණයක් භාවිතා කරමු.
මදුරුවෙක් ඔබේ අත මත පතිත වන බව සිතන්න. මෙම කෘමියාගේ වික්රියාව, එතරම් සැහැල්ලු, අතේ එම ප්රදේශයේ යාන්ත්රික ප්රතිග්රාහක උත්තේජනය කරයි. එම යාන්ත්රික ප්රතිග්රාහක ඔවුන් සම්බන්ධ කර ඇති නියුරෝනය දිගේ පණිවිඩයක් යවයි. නියුරෝනය මොළයට සම්බන්ධ වන අතර එමඟින් පණිවිඩය යවන ලද නිශ්චිත යාන්ත්රික ප්රතිග්රාහකයේ නිශ්චිත ස්ථානයේ ඔබේ ශරීරය ස්පර්ශ කරන බවට පණිවිඩය ලැබේ. මොළය මෙම උපදෙස් සමඟ ක්රියා කරනු ඇත. සමහර විට එය අත්සන හඳුනාගත් අතේ කලාපය දෙස බැලීමට ඇස්වලට කියනු ඇත. ඒවගේම අතේ මදුරුවෙක් ඉන්නවා කියලා ඇස්වලින් මොලේට කිව්වම, මොලේ අතින් කියන්න පුළුවන් ඉක්මනින් ඌව අහකට දාන්න කියලා. යාන්ත්රික ප්රතිග්රාහක ක්රියා කරන්නේ එලෙසයි. පහත ලිපියේ අරමුණ යාන්ත්රික ප්රතිග්රාහකවල ක්රියාකාරී සංවිධානය සහ අණුක නිර්ණායක ප්රදර්ශනය කිරීම මෙන්ම විස්තරාත්මකව සාකච්ඡා කිරීමයි.
ස්පර්ශ සංවේදනය: යාන්ත්රික සංවේදී ප්රතිග්රාහකවල ක්රියාකාරී සංවිධානය සහ අණුක නිර්ණායක
වියුක්ත
චර්ම යාන්ත්රික ප්රතිග්රාහක සමේ විවිධ ස්ථරවල ස්ථානගත කර ඇති අතර එහිදී සැහැල්ලු බුරුසුව, දිගු කිරීම, කම්පනය සහ හානිකර පීඩනය ඇතුළු පුළුල් පරාසයක යාන්ත්රික උත්තේජක හඳුනා ගනී. මෙම උත්තේජක විවිධත්වය විශේෂිත යාන්ත්රික ප්රතිග්රාහක සමූහයක් මගින් සමපාත වන අතර එය විශේෂිත ආකාරයකින් චර්ම විරූපණයට ප්රතිචාර දක්වන අතර මෙම උත්තේජක ඉහළ මොළයේ ව්යුහයන් වෙත යොමු කරයි. යාන්ත්රික ප්රතිග්රාහක සහ ජානමය වශයෙන් ගතහැකි සංවේදී ස්නායු අවසානය පිළිබඳ අධ්යයනයන් ස්පර්ශ සංවේදන යාන්ත්රණයන් අනාවරණය කර ගැනීමට පටන් ගෙන ඇත. මෙම ක්ෂේත්රයේ වැඩ පර්යේෂකයන්ට ස්පර්ශය පිළිබඳ සංජානනයට යටින් පවතින පරිපථ සංවිධානය පිළිබඳ වඩාත් ගැඹුරු අවබෝධයක් ලබා දී ඇත. නව අයන නාලිකා සම්ප්රේෂණ අණු සඳහා අපේක්ෂකයින් ලෙස මතු වී ඇති අතර යාන්ත්රිකව දොරටු ධාරා වල ගුණාංග ස්පර්ශක උත්තේජකවලට අනුවර්තනය වීමේ යාන්ත්රණයන් පිළිබඳ අපගේ අවබෝධය වැඩි දියුණු කළේය. මෙම සමාලෝචනය මගින් රෝම සහ ග්ලැසියර සමෙහි යාන්ත්රික ප්රතිග්රාහකවල ක්රියාකාරී ගුණාංග සහ යාන්ත්රික යෙදවුම් හඳුනා ගන්නා සහ යාන්ත්රික ප්රතිග්රාහක අනුවර්තනය හැඩගස්වන අයන නාලිකාවල ක්රියාකාරී ගුණාංග සංලක්ෂිත කිරීමේ ප්රගතිය ඉස්මතු කරයි.
ප්රධාන වචන: යාන්ත්රික ප්රතිග්රාහක, යාන්ත්රික සංවේදී නාලිකාව, වේදනාව, සම, somatosensory පද්ධතිය, ස්පර්ශය
හැදින්වීම
ස්පර්ශය යනු හානිකර සහ හානිකර යාන්ත්රික උත්තේජක ඇතුළුව සමට බලපාන යාන්ත්රික උත්තේජක හඳුනාගැනීමයි. එය ක්ෂීරපායීන්ගේ සහ මිනිසුන්ගේ පැවැත්මට සහ සංවර්ධනය සඳහා අත්යවශ්ය හැඟීමකි. ඝන ද්රව්ය හා තරල සම හා සම්බන්ධ වීමෙන් මධ්යම ස්නායු පද්ධතියට අවශ්ය තොරතුරු ලබා දෙන අතර එමඟින් පරිසරය ගවේෂණය කිරීමට සහ හඳුනා ගැනීමට ඉඩ ලබා දෙන අතර චලනය හෝ සැලසුම් කළ අත් චලනය ආරම්භ කරයි. ආධුනිකත්වය, සමාජ සම්බන්ධතා සහ ලිංගිකත්වය සඳහා ස්පර්ශය ද ඉතා වැදගත් වේ. බොහෝ ව්යාධි තත්ත්වයන්හිදී එය විකෘති කළ හැකි වුවද (අධිස්රාවය, හයිපෝස්ටේෂියා) ස්පර්ශය අවම වශයෙන් අවදානමට ලක්විය හැකි ඉන්ද්රිය වේ.1-3
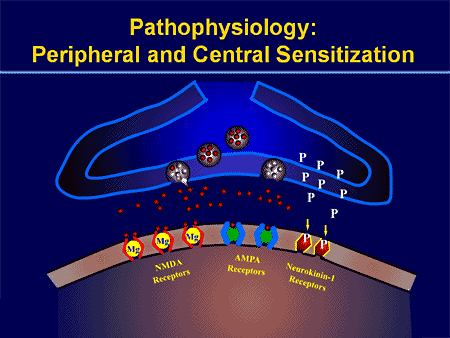
ස්පර්ශ ප්රතිචාර වලට යාන්ත්රික තොරතුරු ඉතා නිවැරදි කේතීකරණයක් ඇතුළත් වේ. චර්ම යාන්ත්රික ප්රතිග්රාහක සමේ විවිධ ස්ථරවල ස්ථානගත කර ඇති අතර එහිදී සැහැල්ලු බුරුසුව, දිගු කිරීම, කම්පනය, හිසකෙස් අපගමනය සහ හානිකර පීඩනය ඇතුළු පුළුල් පරාසයක යාන්ත්රික උත්තේජක හඳුනා ගනී. මෙම උත්තේජක විවිධත්වය විශේෂිත යාන්ත්රික ප්රතිග්රාහක සමූහයක් මගින් සමපාත වන අතර එය විශේෂිත ආකාරයකින් චර්ම විරූපණයට ප්රතිචාර දක්වන අතර මෙම උත්තේජක ඉහළ මොළයේ ව්යුහයන් වෙත යොමු කරයි. සමේ Somatosensory නියුරෝන කාණ්ඩ දෙකකට වැටේ: අඩු එළිපත්ත යාන්ත්රික ප්රතිග්රාහක (LTMRs) නිරපේක්ෂ පීඩනයට ප්රතික්රියා කරයි සහ හානිකර යාන්ත්රික උත්තේජනයට ප්රතිචාර දක්වන ඉහළ එළිපත්ත යාන්ත්රික ප්රතිග්රාහක (HTMRs). LTMR සහ HTMR සෛල සිරුරු පෘෂ්ඨීය මූල ගැන්ග්ලියා (DRG) සහ හිස් කබල සංවේදී ගැන්ග්ලියා (ත්රිකෝණාකාර ගැන්ග්ලියා) තුළ පවතී. LTMRs සහ HTMRs හා සම්බන්ධ ස්නායු කෙඳි A?-, A?- හෝ C-fibers ලෙස වර්ගීකරණය කර ඇත්තේ ඒවායේ ක්රියාකාරී විභව සන්නායක ප්රවේග මත පදනම්වය. C තන්තු මයිලිනීකෘත නොවන අතර අඩුම සන්නායක ප්රවේග (~2 m/s) ඇති අතර A? සහ? තන්තු පිළිවෙළින් අතරමැදි (~12 m/s) සහ වේගවත් (~20 m/s) සන්නායක ප්රවේග ප්රදර්ශනය කරමින් සැහැල්ලුවෙන් හා දැඩි ලෙස මයිලින් කර ඇත. LTMRs තිරසාර යාන්ත්රික උත්තේජකවලට අනුවර්තනය වීමේ අනුපාත අනුව සෙමින් හෝ වේගයෙන් අනුවර්තනය වන ප්රතිචාර (SA- සහ RA-LTMRs) ලෙසද වර්ග කෙරේ. ඔවුන් නවීකරණය කරන චර්ම අන්ත ඉන්ද්රියයන් සහ ඔවුන් කැමති උත්තේජක මගින් ඒවා තවදුරටත් කැපී පෙනේ.
යාන්ත්රික සංඥා හඳුනා ගැනීමට යාන්ත්රික ප්රතිග්රාහකවලට ඇති හැකියාව රඳා පවතින්නේ යාන්ත්රික බලවේග ඉක්මනින් විද්යුත් සංඥා බවට පරිවර්තනය කර ග්රාහක ක්ෂේත්රය වි ධ්රැවීකරණය කරන යාන්ත්රික සම්ප්රේෂක අයන නාලිකා පැවතීම මත ය. ප්රතිග්රාහක විභවය ලෙස හඳුන්වන මෙම ප්රාදේශීය විධ්රැවීකරණය මධ්යම ස්නායු පද්ධතිය දෙසට ව්යාප්ත වන ක්රියාකාරී විභවයන් ජනනය කළ හැක. කෙසේ වෙතත්, යාන්ත්රික සම්ප්රේෂණය සහ යාන්ත්රික බලවේග වලට අනුවර්තනය වීම මැදිහත් වන අණු වල ගුණාංග අපැහැදිලිව පවතී.
මෙම සමාලෝචනයේදී, අපි රෝම සහ දිලිසෙන සමෙහි හානිකර සහ හානිකර ස්පර්ශයේ ක්ෂීරපායී යාන්ත්රික ප්රතිග්රාහක ගුණාංග පිළිබඳ දළ විශ්ලේෂණයක් සපයන්නෙමු. යාන්ත්රික ප්රතිග්රාහක අනුවර්තනය වීමේ යාන්ත්රණය පැහැදිලි කිරීමට උත්සාහ කිරීමේදී යාන්ත්රිකව දොරටු ධාරා වල ගුණාංග පිළිබඳ මෑත කාලීන දැනුම ද අපි සලකා බලමු. අවසාන වශයෙන්, යාන්ත්රික දොරටු ධාරා උත්පාදනය සඳහා වගකිව යුතු අයන නාලිකා සහ ආශ්රිත ප්රෝටීන හඳුනාගැනීමේ මෑතකාලීන ප්රගතිය අපි සමාලෝචනය කරමු.
අහිංසක ස්පර්ශය
රෝම කූප ආශ්රිත LTMRs
රෝම කූප සැහැල්ලු ස්පර්ශය හඳුනා ගන්නා හිසකෙස් පතුවළ නිපදවන කුඩා අවයව නියෝජනය කරයි. කෙස් කළඹ ආශ්රිත කෙඳි, කේශ චලිතයට සහ එහි දිශාවට ප්රතිචාර දක්වන්නේ උත්තේජකය ආරම්භයේදී සහ ඉවත් කිරීමේදී ක්රියාකාරී විභවයන් සහිත දුම්රියන් වෙඩි තැබීමෙනි. ඒවා වේගයෙන් අනුවර්තනය වන ප්රතිග්රාහක වේ.
බළලා හා හාවා. බළලුන් සහ හාවා කබායෙහි, රෝම කූප වර්ග තුනකට බෙදිය හැකිය, පහළ හිසකෙස්, ගාඩ් හිසකෙස් සහ ටයිලොට්රික්ස්. පහතට හිසකෙස් (යටි හිසකෙස්, ලොම්, vellus) 4 කබායෙහි වඩාත්ම බොහෝ, කෙටිම සහ හොඳම හිසකෙස් වේ. ඒවා රැලි සහිත, අවර්ණ වන අතර සමෙහි ඇති පොදු සිදුරකින් හිසකෙස් දෙකේ සිට හතර දක්වා කණ්ඩායම් වශයෙන් මතු වේ. ගාර්ඩ් රෝම (මොනොට්රිච්, ඕවර්හර්ස්, ටොෆෙයාර්) 4 තරමක් වක්ර වී ඇත, එක්කෝ වර්ණක හෝ නොගැලපෙන අතර, ඒවායේ ෆොසිලයේ මුඛයෙන් තනිව මතුවිය. ටයිලෝට්රිච් යනු අඩුම සංඛ්යාවක්, දිගම සහ ඝනතම හිසකෙස් වේ. රෝම කූපයකට සපයන සංවේදී තන්තු සෙබස් ග්රන්ථියට පහළින් පිහිටා ඇති අතර එය A ට ආරෝපණය කර තිබේද? හෝ A?-LTMR තන්තු.5,6
පහළ හිසකෙස් පතුවළට සමීපව, සෙබස් ග්රන්ථියේ මට්ටමට මදක් පහළින් ලැන්සිලේට් පිලෝ-රුෆිනි කෙළවරේ වළල්ල ඇත. මෙම සංවේදී ස්නායු අවසානය කෙස් කලඹ සෑදෙන සම්බන්ධක පටක තුළ කෙස් ගස වටා සර්පිලාකාර මාර්ගයක ස්ථානගත කර ඇත. කෙස් කළඹ තුළ නිදහස් ස්නායු අවසානය ද ඇත, ඒවායින් සමහරක් යාන්ත්රික ප්රතිග්රාහක සාදයි. බොහෝ විට, ටයිලෝට්රිච් ෆොසිලයේ ගෙල ප්රදේශය වටේට ස්පර්ශ කෝපස්කල් (ග්ලැසියර සම බලන්න).
බළලුන්ගේ සහ හාවාගේ ලෝම සහිත සමෙහි ඇති මයිලීන් ස්නායු අවසානයෙහි ගුණ 1930-1970 කාලපරිච්ඡේදය තුළ තීව්ර ලෙස ගවේෂණය කර ඇත (හමාන්, 1995 දී සමාලෝචනය). සහ හාවා, පහළ හිසකෙස් (D වර්ගයේ ප්රතිග්රාහක), ගාඩ් හිසකෙස් (G ප්රතිග්රාහක) සහ Tylotrich හිසකෙස් (T වර්ගයේ ප්රතිග්රාහක) චලනයන්ට අනුරූප වන ප්රතිග්රාහක වර්ග තුනකින් ප්රතිචාර වර්ගීකරණය කර ඇත. RA II නම් පැසීනියානු ප්රතිග්රාහකයට විරුද්ධ වීමෙන් I (RA I) වර්ගයේ වේගයෙන් අනුවර්තනය වන ප්රතිග්රාහකයේ. RA I යාන්ත්රික ප්රතිග්රාහක යාන්ත්රික උත්තේජකයේ ප්රවේගය හඳුනා ගන්නා අතර තියුණු මායිමක් ඇත. ඔවුන් තාප විචලනයන් හඳුනා නොගනී. Burgess et al. pilo-Ruffini අවසානය උත්තේජනය කිරීම නිසා සමට හෝ හිසකෙස් කිහිපයක චලනයට ප්රශස්ත ලෙස ප්රතිචාර දක්වන වේගයෙන් අනුවර්තනය වන ක්ෂේත්ර ප්රතිග්රාහකයක් ද විස්තර කර ඇත. C ෆයිබර් ක්රියාකාරිත්වයට කෙස් කළඹේ ප්රතිචාර කිසිවක් ආරෝපණය කර නැත.8
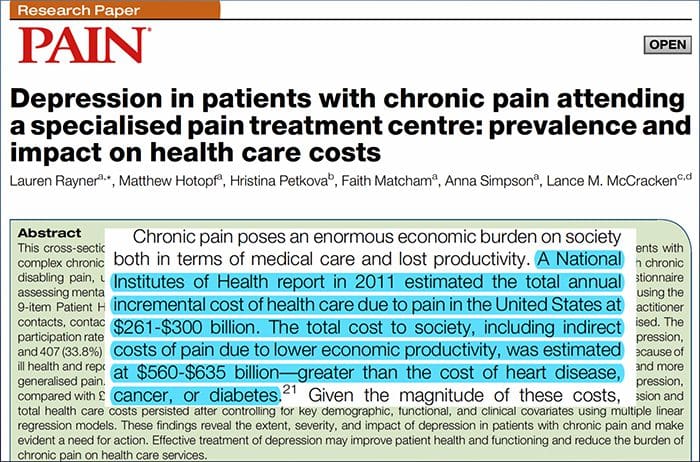
මීයන්. මීයන්ගේ පෘෂ්ඨීය රෝම සහිත සමෙහි, ප්රධාන රෝම කූප වර්ග තුනක් විස්තර කර ඇත: සිග්සැග් (72% පමණ), awl/auchene (23% පමණ) සහ ගාඩ් හෝ ටයිලෝට්රිච් (5% පමණ).11-14 Zigzag සහ Awl/ auchenne රෝම කූප තුනී සහ කෙටි කෙස් ගස් නිපදවන අතර එක් sebaceous ග්රන්ථියක් සමඟ සම්බන්ධ වේ. ගාර්ඩ් හෝ ටයිලෝට්රිච් කෙස් යනු කෙස් කළඹේ දිගම වේ. ඔවුන් sebaceous ග්රන්ථි දෙකක් සමඟ සම්බන්ධ විශාල හිසකෙස් බල්බයක් මගින් සංලක්ෂිත වේ. ගාඩ් සහ awl/auchene හිසකෙස් පුනරාවර්තන, නිතිපතා පරතරයකින් යුත් රටාවකට සකසා ඇති අතර සිග්සැග් හිසකෙස් විශාල රෝම කූප වර්ග දෙක අවට සම ප්රදේශ ඝන ලෙස ජනාකීර්ණ කරයි [රූපය. 1 (A1, A2 සහ A3)].
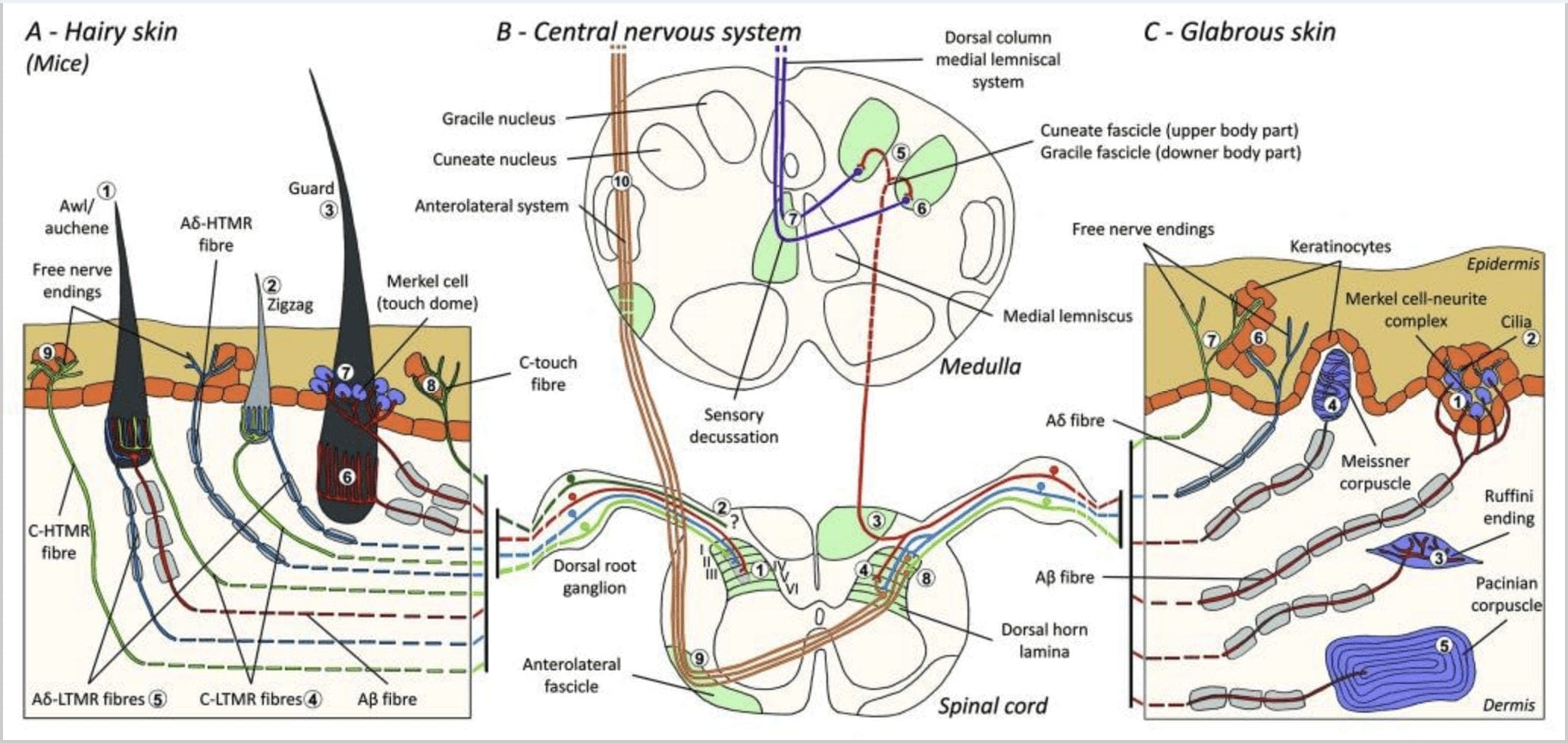
රූපය 1. චර්ම යාන්ත්රික ප්රතිග්රාහක සංවිධානය සහ ප්රක්ෂේපණ. හිසකෙස් ඇති සමෙහි, සැහැල්ලු බුරුසුව සහ ස්පර්ශය ප්රධාන වශයෙන් අනාවරණය වන්නේ කෙස් කළඹ වටා ඇති නවෝත්පාදනය මගිනි: awl/auchenne (A1), zigzag (A2) සහ guard (A3). Awl/auchene හිසකෙස් C-LTMR lanceolate endings (A4), A?-LTMR සහ A මගින් තුන් ගුණයකින් නවීකරණය කර ඇත. වේගයෙන් අනුවර්තනය වීම-LTMR (A6). සිග්සැග් කෙස් කළඹ යනු කෙටි කෙස් පතුවළ වන අතර ඒවා C-LTMR (A4) සහ A යන දෙකින්ම නවීකරණය කර තිබේද? -LTMR lanceolate endings (A5). දිගම ආරක්ෂක රෝම කූප නවීකරණය කර ඇත්තේ A විසින්? වේගයෙන් අනුවර්තනය වීම-LTMR කල්පවත්නා ලැන්සිලේට් අවසානය (A6) සහ A සමඟ සම්බන්ධ වී තිබේද? සෙමින් අනුවර්තනය වීම-LTMR ස්පර්ශ dome endings (A7). මෙම සියලුම තන්තු වල කේන්ද්රීය ප්රක්ෂේපණය සුෂුම්නාව පෘෂ්ඨීය අං වල වෙනස්, නමුත් අර්ධ වශයෙන් අතිච්ඡාදනය වන ලැමිනේ වලින් අවසන් වේ (C-LTMR lamina II, A?-LTMR lamina III සහ A?-LTMR lamina IV සහ V). එකම හෝ යාබද රෝම කූප නවීකරණය කරන LTMR හි ප්රක්ෂේපණ සුෂුම්නාව පෘෂ්ඨීය අං (අළු පැහැයෙන් B1) හි පටු තීරුවක් සෑදීමට පෙළගස්වා ඇත. ලෝම සහිත සමෙහි පමණක්, C-තන්තු රහිත අන්තයේ උප ජනගහනයක් එපීඩර්මිස් නවීකරණය කරන අතර ප්රසන්න ස්පර්ශයට ප්රතිචාර දක්වයි (A8). මෙම C-ස්පර්ශ තන්තු හානිකර ස්පර්ශයට ප්රතිචාර නොදක්වන අතර ඒවායේ ගමන් මාර්ගය තවමත් හඳුනාගෙන නොමැත (B2). දිලිසෙන සමෙහි, අහිංසක ස්පර්ශය LTMR වර්ග හතරකින් මැදිහත් වේ. මර්කෙල් සෛල-නියුරයිට් සංකීර්ණය එපීඩර්මිස් (C1) හි බාසල් ස්ථරයේ ඇත. මෙම යාන්ත්රික ප්රතිග්රාහකය බොහෝ මර්කල් සෛල අතර සැකැස්මකින් සහ තනි A එකකින් විශාල වූ ස්නායු අග්රයකින් සමන්විතද? තන්තු. මර්කෙල් සෛල keratinocytes (C2) සම්බන්ධ කරන ක්රියාවලීන් වැනි ඇඟිල්ලක් පෙන්වයි. රුෆිනි අවසානය ඩර්මිස් තුළ ස්ථානගත කර ඇත. එය A හා සම්බන්ධ තුනී සුරුට්ටු හැඩැති සංවෘත සංවේදී අවසානයක්ද? තන්තු (C3). A ට සම්බන්ධ වූ Meissner corpuscle? ස්නායු අවසානය සහ සමේ පැපිලිය තුළ පිහිටා ඇත. මෙම සංවෘත යාන්ත්රික ප්රතිග්රාහකය සමන්විත වන්නේ සම්බන්ධක පටක (C4) වලින් වට වූ තිරස් ලැමිලේ ලෙස සකසා ඇති ඇසුරුම් කරන ලද ආධාරක සෛල වලින්ය. Pacinian corpuscle යනු ගැඹුරු යාන්ත්රික ප්රතිග්රාහකයයි. තනි A? සංකේන්ද්රීය ලැමිලේ වලින් සෑදූ මෙම විශාල ඩිම්බ කෝෂ වල මධ්යයේ මයිලින් නොකළ ස්නායු අවසානය අවසන් වේ. සුෂුම්නාව තුළ ඇති මෙම A?-LTMR තන්තු වල ප්රක්ෂේපන ශාඛා දෙකකට බෙදා ඇත. ප්රධාන මධ්යම ශාඛාව (B3) කොඳු ඇට පෙළේ ඉප්සිලේටරල් පෘෂ්ඨීය ප්රදේශයේ ඉහළට නැඟී, ප්රාථමික අනුග්රාහකයින් ඔවුන්ගේ පළමු උපාගම (B5) සිදු කරන මෙඩුල්ලා මට්ටමින් උල් හෝ ග්රැසිල් ෆැසිකල් (B6) සාදයි. ද්විතියික නියුරෝන මගින් මොළයේ කඳ හරහා මධ්ය මොළයට, විශේෂයෙන් තලමස් වෙත නැඟී එන මධ්ය ලෙම්නිස්කස් මත පත්රිකාවක් සෑදීමට සංවේදී නියුරෝන (B7) සිදු කරයි. LTMR හි ද්විතියික ශාඛාව ලැමිනා II, IV, V හි පෘෂ්ඨීය අං වලින් අවසන් වන අතර වේදනාව සම්ප්රේෂණයට (B4) බාධා කරයි. ලෝම (A9) සහ දිලිසෙන සම (C7) යන දෙකෙහිම එපීඩර්මිස් හි නිදහස් ස්නායු අවසානය මගින් හානිකර ස්පර්ශය අනාවරණය වේ. මෙම යාන්ත්රික ප්රතිග්රාහක යනු අසල්වැසි කෙරටිනොසයිට් (C6) සමඟ සමීප සම්බන්ධතා ඇති A?-HTMR සහ C-HTMR වල අවසානයයි. A?-hTMR ලැමිනා I සහ V හි අවසන් වේ; C-HTMR ලැමිනා I සහ II (B8) තුළ අවසන් වේ. සුෂුම්නාව පෘෂ්ඨීය අං මට්ටමේ දී, ප්රාථමික අනුග්රාහක HTMRs ද්විතියික නියුරෝන සමඟ උපාගම සාදයි, එය මධ්ය රේඛාව හරහා ගොස් ප්රති-පාර්ශවීය ෆැසිසිකලයේ (B9, B10) ඉහළ මොළයේ ව්යුහයට නැඟේ.
මෑතදී, Ginty සහ සහකාරකයින් මීයන් තුළ LTMR වල පර්යන්ත සහ මධ්යම අක්ෂීය අවසානයන් සංවිධානය කිරීම දෘශ්යමාන කිරීම සඳහා අණුක-ජාන ලේබල් කිරීම සහ somatotopic ප්රතිගාමී ලුහුබැඳීමේ ප්රවේශයන් වල එකතුවක් භාවිතා කළහ.15 ඔවුන්ගේ සොයාගැනීම් සංකීර්ණ ස්පර්ශක උත්තේජකයේ තනි ලක්ෂණ ඇති ආකෘතියකට සහාය වේ. රෝම කූප වර්ග තුනෙන් නිස්සාරණය කර A?-, A?- සහ C- තන්තු වල අද්විතීය සංයෝජනවල ක්රියාකාරකම් හරහා පෘෂ්ඨීය අං වෙත සම්ප්රේෂණය වේ.
ටයිරොසීන් හයිඩ්රොක්සිලේස් පොසිටිව් (TH+) DRG නියුරෝන වල ජානමය ලේබල් කිරීම මගින් පෙප්ටයිඩර්ජික් නොවන, කුඩා-විෂ්කම්භය සංවේදී නියුරෝන ජනගහනයක් සංලක්ෂිත වන අතර සමේ C-LTMR පර්යන්ත අවසානයන් දෘශ්යමාන කිරීමට ඉඩ සලසයි. පුදුමයට කරුණක් නම්, තනි තනි C-LTMR වල අක්ෂීය ශාඛා, සිග්සැග් (අවසාන වලින් 80%) සහ awl/auchene (අවසාන වලින් 20%) සමඟ සමීපව සම්බන්ධ වන කල්පවත්නා ලැන්සිලේට් අවසානයන් වෘක්ෂලතා කර ඇති බව සොයා ගන්නා ලදී, නමුත් tylotrich රෝම කූප නොවේ [Fig. 1 (A4)]. Longitudinal lanceolate endings A?-LTMRs වලට පමණක් අයත් යැයි බොහෝ කලක සිට සිතූ අතර එබැවින් C-LTMR වල අවසානය කල්පවත්නා lanceolate endings සාදනු ඇතැයි අනපේක්ෂිත විය. myelinated mechanoreceptors [පය. 15 (C2)].
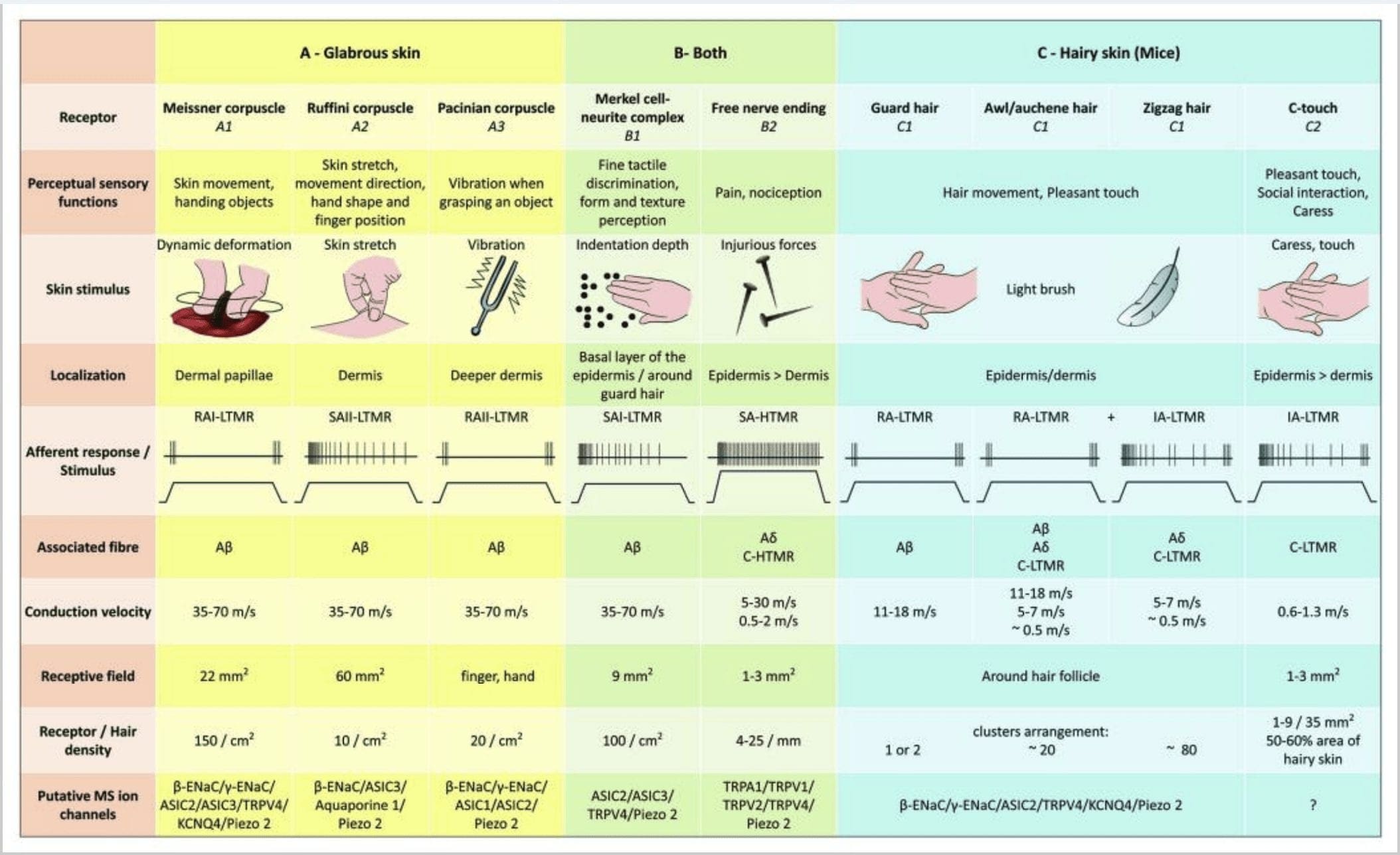
රූපය 2. ක්ෂීරපායීන්ගේ ස්පර්ශ ප්රතිග්රාහක: චර්ම ස්පර්ශක ප්රතිග්රාහක, දිලිසෙන සහ රෝම සහිත සමෙහි අඩු යාන්ත්රික එළිපත්ත (LTMRs) සහ ඉහළ යාන්ත්රික එළිපත්ත ප්රතිග්රාහක (HTMRs) මගින් සහාය දක්වන හානිකර ස්පර්ශය සහිත බහු ප්රතිග්රාහක මගින් සහය දක්වන අහිංසක ස්පර්ශයට වෙනස් වේ. ඒවා ප්රධාන වශයෙන් එපීඩර්මිස් වලින් අවසන් වන ස්නායු රහිත අවසානය සාදයි. (A) දිලිසෙන සම. A1: Meissner corpuscles සමේ චලනය සහ අතේ ඇති වස්තුව ලිස්සා යාම හඳුනා ගනී. වස්තුව සහ දක්ෂතාවය භාර දීම සඳහා ඒවා වැදගත් වේ. ප්රතිග්රාහක වේගයෙන් උත්තේජකයට අනුවර්තනය වේ, A සමඟ සම්බන්ධ වී තිබේද? තන්තු සහ කලාතුරකින් C තන්තු දක්වා විශාල ප්රතිග්රාහක ක්ෂේත්රයක් ඇත. A2: Ruffini corpuscles සමේ දිගුව හඳුනා ගන්නා අතර ඇඟිලි පිහිටීම සහ වස්තුව ලබා දීම සඳහා වැදගත් වේ. ප්රතිග්රාහකය සෙමෙන් උත්තේජකයට අනුවර්තනය වන අතර උත්තේජකය යොදන තාක් කල් ක්රියාකාරකම් පවත්වා ගනී. ප්රතිග්රාහක A සමඟ සම්බන්ධ වී තිබේද? තන්තු සහ විශාල ප්රතිග්රාහක ක්ෂේත්රයක් ඇත. A3: Pacinian corpuscles dermis තුළ ගැඹුරු වන අතර කම්පනය හඳුනා ගනී. ප්රතිග්රාහක A සමඟ සම්බන්ධ වී තිබේද? කෙඳි; ඔවුන් ඉක්මනින් උත්තේජකයට අනුවර්තනය වන අතර විශාලතම ප්රතිග්රාහක ක්ෂේත්රය ඇත. (B) සම්පූර්ණ සම. B1: මර්කෙල්-සෛල සංකීර්ණ දිලිසෙන සම සහ හිසකෙස් අවට යන දෙකෙහිම පවතී. ඒවා අතේ ඝන ලෙස ප්රකාශ වී ඇති අතර කරුණු දෙකක් අතර වයනය සංජානනය සහ හොඳම වෙනස්කම් කිරීම සඳහා වැදගත් වේ. ඇඟිලි නිරවද්යතාව සඳහා ඔවුන් වගකිව යුතුය. ප්රතිග්රාහක A සමඟ සම්බන්ධ වී තිබේද? කෙඳි; ඔවුන් සෙමින් උත්තේජනයට අනුවර්තනය වන අතර කෙටි ප්රතිග්රාහක ක්ෂේත්රයක් ඇත. B2: උත්තේජකයට ඉතා මන්දගාමී අනුවර්තනයක් සහිත හානිකර ස්පර්ශ HTMRs, එනම්, nociceptive උත්තේජකය යොදන තාක් සක්රීය වේ. A හි නිදහස් ස්නායු අවසානය මගින් ඒවා සෑදී ඇත? සහ keratinocytes හා සම්බන්ධ C-තන්තු. (C) හිසකෙස් සහිත සම. C1: කෙස් කළඹ විවිධ හිසකෙස් වර්ග සමඟ සම්බන්ධ වේ. මීයන් තුළ ගාර්ඩ් හිසකෙස් දිගු හා විරල ලෙස ප්රකාශිත එකක් වන අතර, awl/auchenne මධ්යම ප්රමාණයේ වන අතර සිග්සැග් කුඩාම සහ වඩාත් ඝන ලෙස ප්රකාශිත හිසකෙස් වේ. ඔවුන් A සමඟ සම්බන්ධ වී තිබේද? කෙඳි නමුත් A වෙතද? සහ awl/auchenne සහ zizag හිසකෙස් සඳහා C-LTMRs කෙඳි. ඔවුන් සෙනෙහස අතරතුර ප්රසන්න ස්පර්ශය ඇතුළුව හිසකෙස් චලනය හඳුනා ගනී. ඔවුන් උත්තේජක සඳහා වේගයෙන් හෝ අතරමැදි චාලක සමග අනුගත වේ. C2: C-ස්පර්ශ ස්නායු අවසානය අඩු යාන්ත්රික එළිපත්තකින් සංලක්ෂිත නිදහස් අවසානයක් සහිත C තන්තු පර්යන්තයේ උප වර්ගයකට අනුරූප වේ. ඔවුන් සෙනෙහස නිසා ඇතිවන ප්රසන්න සංවේදනය සඳහා කේතනය කළ යුතුය. ඔවුන් උත්තේජනයට මධ්යස්ථව අනුගත වන අතර කෙටි ප්රතිග්රාහක ක්ෂේත්රයක් ඇත. විවිධ ස්පර්ශක ප්රතිග්රාහකවල ප්රකාශිත යාන්ත්රික සංවේදී (MS) අයන නාලිකා මූලික දත්තවලට අනුව දක්වනු ලබන අතර ඇගයීම යටතේ පවතින කල්පිතය සාරාංශ කරයි.
හදුනාගත් දෙවන ප්රධාන ජනගහණයක් Awl/Auchenne සහ zigzag follicles හි A?-LTMR අවසානය ගැන සැලකිලිමත් වන අතර, බළලුන් සහ හාවා තුළ පුළුල් ලෙස අධ්යයනය කරන ලද Down hair follicle සමඟ සංසන්දනය කළ යුතුය. Ginty සහ සහයෝගිතාකරුවන් පෙන්වා දුන්නේ TrkB මධ්යම-විෂ්කම්භය DRG නියුරෝනවල උප කුලකයක ඉහළ මට්ටමකින් ප්රකාශ වන බවයි. ලේබල් කරන ලද තන්තු වල ex vivo සම-ස්නායු සකස් කිරීම භාවිතා කරන අන්තර් සෛලීය පටිගත කිරීම් මගින් හෙළි වූයේ ඒවා කලින් බළලුන් සහ හාවා තුළ අධ්යයනය කරන ලද තන්තු වල භෞතික විද්යාත්මක ගුණාංග ප්රදර්ශනය කරන බවයි: අතිවිශිෂ්ට යාන්ත්රික සංවේදීතාව (Von Frey threshold <0.07 mN), suprathreshold උත්තේජක සඳහා ප්රතිචාර වේගයෙන් අනුවර්තනය වීම, අතරමැදි. ප්රවේග (5.8 ± 0.9 m/s) සහ පටු නොගැලපෙන සෝමා කරල්. 15 (A1)].
අවසාන වශයෙන්, ඔවුන් පෙන්වා දුන්නේ A වේගයෙන් අනුවර්තනය වීමේ පර්යන්ත අවසානය බවයි? LTMRs ආරක්ෂක (හෝ tylotrich) සහ awl/auchene රෝම කූප සමඟ සම්බන්ධ කල්පවත්නා ලැන්සිලේට් අවසානය සාදයි [රූපය. 1 (A6)].15 මීට අමතරව, A ට සම්බන්ධ වූ ටච් ඩෝම් සාදමින් මර්කල් සෛල සංකීර්ණයක් සමඟ ආරක්ෂක හිසකෙස් ද සම්බන්ධ වේ? LTMR සෙමින් අනුවර්තනය වීම [රූපය. 1 (A7)].
සාරාංශයක් ලෙස, සියලුම සිග්සැග් රෝම කූප C-LTMR සහ A?-LTMR lanceolate endings යන දෙකින්ම නවීකරණය කර ඇත; A විසින් awl/auchene හිසකෙස් තුන් ගුණයකින් නවීකරණය කර තිබේද? වේගයෙන් අනුවර්තනය වීම-LTMR, A?-LTMR සහ C-LTMR lanceolate endings; ආරක්ෂක රෝම කූප A මගින් නවීකරණය කරයිද? වේගයෙන් අනුවර්තනය වීම-LTMR කල්පවත්නා ලැන්සිලේට් අවසානය සහ A සමඟ අන්තර්ක්රියා කරයිද? සෙමින් අනුවර්තනය වීම-LTMR ස්පර්ශ dome endings. මේ අනුව, සෑම මූසික රෝම කූපයකටම ස්නායු භෞතික විද්යාත්මකව වෙනස් යාන්ත්රික සංවේදී අවසාන අවයව වලට අනුරූප වන LTMR අවසානයන්හි අද්විතීය සහ වෙනස් නොවන සංයෝජන ලැබේ. මෙම හිසකෙස් වර්ග තුනේ පුනරාවර්තන සැකැස්ම සැලකිල්ලට ගනිමින්, Ginty සහ සහකාරකයින් යෝජනා කරන්නේ හිසකෙස් සහිත සම යනු පර්යන්ත ඒකකයේ පුනරාවර්තන පුනරාවර්තනයකින් සමන්විත වන බවයි ~1 අතරමැදි සිග්සැග් හිසකෙස් [පය. 2 (C20)].
කොඳු ඇට පෙළේ ප්රක්ෂේපණය. A හි කේන්ද්රීය ප්රක්ෂේපණය? ශීඝ්රයෙන් අනුවර්තනය වන-LTMRs, A?-LTMRs සහ C-LTMRs සුසුම්නාවේ පෘෂ්ඨීය අංවල වෙනස්, නමුත් අර්ධ වශයෙන් අතිච්ඡාදනය වන ලැමිනේ (II, III, IV) වලින් අවසන් වේ. මීට අමතරව, පර්යන්ත LTMR ඒකකයක් තුළ එකම හෝ යාබද රෝම කූප නවීකරණය කරන LTMR වල මධ්යම පර්යන්ත සුෂුම්නාව පෘෂ්ඨීය අං තුළ පටු LTMR තීරුවක් සෑදීමට පෙළගස්වා ඇත [රූපය. 1 (B1)]. මේ අනුව, පෘෂ්ඨීය අං තුළ ඇති සෝමාටෝටොපිකල් ලෙස සංවිධානය වූ ප්රාථමික සංවේදක අනුක්රමික අවසානයන්හි කුඤ්ඤයක් හෝ තීරුවක් එකම පර්යන්ත ඒකකය නවීකරණය කරන සහ යාන්ත්රික හඳුනා ගන්නා A?-, A?- සහ C-LTMR වල මධ්ය ප්රක්ෂේපණවල පෙළගැස්ම නියෝජනය කරන බව පෙනේ. එකම කුඩා රෝම කූප මත ක්රියා කරන උත්තේජක. කඳේ සහ අත් පා වල ගාඩ්, awl/auchene සහ zigzag රෝම සංඛ්යාව සහ එක් එක් LTMR උපවර්ගයේ සංඛ්යා මත පදනම්ව, Ginty සහ සහකාරකයන් ඇස්තමේන්තු කරන්නේ මූසික පෘෂ්ඨීය අං වල LTMR තීරු 2,000-4,000 ක් ඇති අතර එය පර්යන්ත ගණනට අනුරූප වේ. LTMR ඒකක.15
තවද, LTMR උප වර්ගවල අක්සෝන එකිනෙකට සමීපව සම්බන්ධ වී ඇති අතර, එම රෝම කූපයම නවීකරණය කරන ප්රක්ෂේපන සහ අන්තර් සංඛ්යාංක ලැන්සිලේට් අවසානයන් ඇත. මීට අමතරව, කෙස් කළඹ වර්ග තුන විවිධ හැඩයන්, ප්රමාණයන් සහ සෛලීය සංයුතීන් ප්රදර්ශනය කරන බැවින්, ඒවාට වෙනස් අපගමනය හෝ කම්පන සුසර කිරීමේ ගුණ තිබිය හැකිය. මෙම සොයාගැනීම් බළලුන්ගේ සහ හාවාගේ සම්භාව්ය ස්නායු භෞතික විද්යාත්මක මිනුම්වලට අනුකූල වන අතර එයින් පෙන්නුම් කරන්නේ A? RA-LTMRs සහ A?-LTMRs එකිනෙකට වෙනස් රෝම කූප වර්ගවල අපගමනය මගින් වෙනස් ලෙස සක්රිය කළ හැක.16,17
අවසාන වශයෙන්, හිසකෙස් සහිත සමෙහි ස්පර්ශය සංකලනය වේ: (1) රෝම කූප වර්ග තුනේ සාපේක්ෂ සංඛ්යා, අද්විතීය අවකාශීය ව්යාප්තිය සහ වෙනස් රූප විද්යාත්මක සහ අපගමන ගුණාංග; (2) එක් එක් රෝම කූප වර්ග තුනට සම්බන්ධ LTMR උපවර්ග අවසානයෙහි අද්විතීය සංයෝජන; සහ (3) හිසකෙස් සහිත සම යාන්ත්රික සංවේදක පද්ධතියට නිස්සාරණය කිරීමට සහ CNS වෙත ලබා දීමට හැකි වන පරිදි කෙස් කළඹ ආශ්රිත LTMR වල ප්රධාන පන්ති හතරේ වෙනස් සංවේදීතාවන්, සන්නායක ප්රවේගයන්, ස්පයික් දුම්රිය රටා සහ අනුවර්තන ගුණාංග ස්පර්ශ කරන්න.
නිදහස් ස්නායු අවසානය LTMRs
සාමාන්යයෙන්, සමෙහි ඇති C-තන්තු රහිත අවසානය HTMR වේ, නමුත් C-තන්තු වල උප ජනගහනයක් හානිකර ස්පර්ශයට ප්රතිචාර නොදක්වයි. මෙම ස්පර්ශක C-ෆයිබර් (CT) අනුකුලිකයන්ගේ උප කුලකය මිනිසුන්ගේ සහ ක්ෂීරපායීන්ගේ රෝම සහිත නමුත් ග්ලැසියර නොවන සමෙහි පවතින වෙනස් නොවන මයිලිනේටඩ්, අඩු එළිපත්ත යාන්ත්රික ප්රතිග්රාහක ඒකක නියෝජනය කරයි [රූපය. 1 (A8)].18,19 CTs සාමාන්යයෙන් ශරීර ස්පර්ශයේදී ප්රසන්න ස්පර්ශ උත්තේජනයක් පිළිබඳ සංජානනය සමඟ සම්බන්ධ වේ.20,21
CT afferents 0.3–2.5 mN පරාසයේ indentation Force වලට ප්රතිචාර දක්වන අතර ඒ අනුව බොහෝ A තරම් සමේ විරූපණයට සංවේදීද? afferents.19 සෙමින් හා වේගයෙන් අනුවර්තනය වන myelinated mechanoreceptors සමඟ සංසන්දනය කිරීමේදී CT afferents හි අනුවර්තන ලක්ෂණ අතරමැදි වේ. මානව CT afferents හි ප්රතිග්රාහක ක්ෂේත්ර දළ වශයෙන් වටකුරු හෝ ඕවලාකාර හැඩයෙන් යුක්ත වේ. ක්ෂේත්රය 35 mm2.22 දක්වා වූ ප්රදේශයක් පුරා බෙදා හරින ලද කුඩා ප්රතිචාරාත්මක ලප එක සිට නවය දක්වා සමන්විත වේ. මූසික සමලිංගික ප්රතිග්රාහක රෝම සමෙහි ප්රදේශයෙන් 50-60% පමණ ආවරණය වන පරිදි අඛණ්ඩ පැච් රටාවකට සංවිධානය කර ඇත [රූපය. 2 (C2)].23
මයිලිනේටඩ් ස්පර්ශක අනුකාරක නොමැති රෝගීන්ගේ සාක්ෂි වලින් පෙන්නුම් කරන්නේ CT තන්තු වල සංඥා කිරීම පරිවාරක බාහිකය සක්රීය කරන බවයි. මෙම පද්ධතිය ස්පර්ශයේ වෙනස් කොට සැලකීමේ අංගයන් කේතනය කිරීමේදී දුර්වල බැවින්, නමුත් මන්දගාමී, මෘදු ස්පර්ශ කේතනය කිරීමට හොඳින් ගැලපෙන බැවින්, හිසකෙස් සහිත සමෙහි ඇති CT තන්තු ස්පර්ශයේ ප්රසන්න සහ සමාජීය වශයෙන් අදාළ අංශ සැකසීමේ පද්ධතියක කොටසක් විය හැකිය.24 CT තන්තු සක්රීය කිරීමද විය හැක. වේදනාව නිවාරණය කිරීමේ කාර්යභාරයක් ඇති අතර දැවිල්ල හෝ කම්පනය C-fiber LTMR මගින් ප්රසන්න ස්පර්ශයේ සිට වේදනාව දක්වා සංවේදනය වෙනස් කළ හැකි බව මෑතකදී යෝජනා කර ඇත.25,26
CT-afferents ගමන් කරන්නේ කුමන මාර්ගයද යන්න තවමත් නොදනී [රූපය. 1 (B2)], නමුත් ස්පිනොතලමික් ප්රක්ෂේපණ සෛල සඳහා අඩු-ඉදිරිපත් වූ ස්පර්ශක යෙදවුම් ලේඛනගත කර ඇත, 27 chordotomy ක්රියා පටිපාටිවලින් පසු මෙම මාර්ග විනාශ වීමෙන් පසු මිනිස් රෝගීන්ගේ ස්පර්ශ හඳුනාගැනීමේ සියුම්, ප්රතිවිරුද්ධ ඌනතාවයන් පිළිබඳ වාර්තා සඳහා විශ්වාසය ලබා දෙයි.
Glabrous Skin හි LTMRs
මර්කල් සෛල-නියුරයිට් සංකීර්ණ සහ ස්පර්ශ ගෝලාකාර. මර්කෙල් (1875) යනු උපකල්පිත ඇෆෙරන්ට් ස්නායු තන්තු සමඟ සම්බන්ධතා ඇති කර ගනිමින් විශාල ලෝබියුලේටඩ් න්යෂ්ටි සහිත එපීඩර්මල් සෛල පොකුරු පිළිබඳ හිස්ටෝල් විද්යාත්මක විස්තරයක් ලබා දුන් ප්රථමයා විය. ඔහු උපකල්පනය කළේ ඔවුන් ඒවා Tastzellen (ස්පර්ශක සෛල) ලෙස හැඳින්වීමෙන් ස්පර්ශය පිළිබඳ හැඟීමට යටත් වූ බවයි. මිනිසුන් තුළ, මර්කල් සෛල නියුරයිට් සංකීර්ණ සමේ ස්පර්ශ සංවේදී ප්රදේශවල පොහොසත් වන අතර ඒවා ඇඟිලි, තොල් සහ ලිංගික අවයවවල එපීඩර්මිස් හි බාසල් ස්ථරයේ දක්නට ලැබේ. ඒවා අඩු ඝනත්වයකින් යුත් කෙස් සහිත සමෙහි ද පවතී. මර්කෙල් සෛල නියුරයිට් සංකීර්ණය සමන්විත වන්නේ තනි මයිලිනේටඩ් A වලින් විශාල වූ ස්නායු අග්රයකට ආසන්නව ඇති මර්කෙල් සෛලයකින්ද? තන්තු [පය. 1 (C1)] (හලාටා සහ සහයෝගිතාකරුවන් තුළ සමාලෝචනය).29 අපිචර්ම පැත්තේ මර්කල් සෛලය අසල්වැසි කෙරටිනොසයිට් අතර විහිදෙන ඇඟිලි වැනි ක්රියාවලීන් ප්රදර්ශනය කරයි [රූපය. 1 (C2)]. මර්කෙල් සෛල යනු කෙරටිනොසයිට්-ව්යුත්පන්න එපීඩර්මල් සෛල වේ.30,31 ටච් ඩෝම් යන යෙදුම හඳුන්වා දෙනු ලැබුවේ බළලුන්ගේ පෙර පාදයේ රෝම සමෙහි ඇති මර්කල් සෛල සංකීර්ණවල විශාල සාන්ද්රණය නම් කිරීමට ය. ටච් ඩෝම් එකක තනි A?-ෆයිබර් එකකින් නවීකරණය කරන ලද මර්කල් සෛල 150ක් දක්වා තිබිය හැකි අතර A?-fibers, A හැර මිනිසුන් තුළද? සහ C-තන්තු ද නිතිපතා පැවතුනි.32-34
මර්කෙල් සෙලෝනියුරයිට් සංකීර්ණ උත්තේජනය කිරීමේ ප්රතිඵලය වන්නේ තියුණු මායිම් සහිත විරාම ග්රාහක ක්ෂේත්රවලින් හටගන්නා I (SA I) ප්රතිචාර සෙමින් අනුවර්තනය වීමයි. ස්වයංසිද්ධ විසර්ජනයක් නොමැත. මෙම සංකීර්ණ සමේ ඉන්ඩෙන්ටේෂන් ගැඹුරට ප්රතිචාර දක්වන අතර චර්ම යාන්ත්රික ප්රතිග්රාහකවල ඉහළම අවකාශීය විභේදනය (0.5 මි.මී.) ඇත. ඔවුන් ස්පර්ශක උත්තේජකවල නිරවද්ය අවකාශීය රූපයක් සම්ප්රේෂණය කරන අතර හැඩය සහ වයනය වෙනස් කිරීම සඳහා වගකිව යුතු යැයි යෝජනා කෙරේ [රූපය. 2 (B1)]. මර්කල් සෛල වලින් තොර මීයන්ට තම උඩු රැවුල භාවිතා කරන අතරතුර ඔවුන්ගේ පාදවලින් වයනය සහිත පෘෂ්ඨයන් හඳුනාගත නොහැක.35
මර්කල් සෛලය, සංවේදක නියුරෝනය හෝ දෙකම යාන්ත්රික සම්ප්රේෂණයේ ස්ථාන ද යන්න තවමත් විවාදයට තුඩු දෙන කරුණකි. මීයන් තුළ, මර්කෙල් සෛලවල ෆොටෝටොක්සික් විනාශය SA I ප්රතිචාරය අහෝසි කරයි. 36 ජානමය වශයෙන් යටපත් කරන ලද-මර්කෙල් සෛල සහිත මීයන් තුළ, ex vivo සම/ස්නායු සැකසීමේදී වාර්තා වූ SA I ප්රතිචාරය සම්පූර්ණයෙන්ම අතුරුදහන් වී, මර්කෙල්ගේ නිසි කේතනය සඳහා මර්කෙල් සෛල අවශ්ය බව පෙන්නුම් කරයි. ප්රතිග්රාහක ප්රතිචාර.37 කෙසේ වෙතත්, මෝටර් ක්රියා කරන පීඩනය මගින් සංස්කෘතියේ හුදකලා වූ මර්කෙල් සෛල යාන්ත්රික උත්තේජනය යාන්ත්රිකව දොරටු ධාරා ජනනය නොකරයි. මර්කල් සෛල ඇඟිලි වැනි ක්රියාවලීන් සමේ විරූපණය සහ එපීඩර්මිස් සෛල චලනය සමඟ ගමන් කළ හැකි අතර මෙය යාන්ත්රික සම්ප්රේෂණයේ පළමු පියවර විය හැකිය. පැහැදිලිවම, මර්කෙල් සෛලවල යාන්ත්රික සංවේදීතාව අධ්යයනය කිරීමට අවශ්ය කොන්දේසි තවම ස්ථාපිත කර නොමැත.
රුෆිනි අවසානය. Ruffini endings යනු A හා සම්බන්ධ වූ තුනී සුරුට්ටු හැඩැති සංවෘත සංවේදී අවසානයද? ස්නායු අවසානය. රූෆිනි අවසානය යනු චර්ම කොලජන් කෙඳි දිගේ සකස් කර ඇති කුඩා සම්බන්ධක පටක සිලින්ඩර වන අතර ඒවා විෂ්කම්භය 4-6 m මයිලිනේටඩ් ස්නායු තන්තු එකකින් හෝ තුනකින් සපයනු ලැබේ. ඩර්මිස් හි විවිධ දිශානතියේ සිලින්ඩර තුනක් දක්වා එක් ප්රතිග්රාහකයක් සෑදීමට ඒකාබද්ධ විය හැකිය [රූපය. 1 (C3)]. ව්යුහාත්මකව, Ruffini අවසානය ගොල්ගි කණ්ඩරාවන්ට සමාන වේ. ඒවා ඩර්මිස් තුළ පුළුල් ලෙස ප්රකාශ වන අතර සෙමින් අනුවර්තනය වන II (SA II) චර්ම යාන්ත්රික ප්රතිග්රාහක ලෙස හඳුනාගෙන ඇත. ස්වයංසිද්ධ ස්නායු ක්රියාකාරිත්වයේ පසුබිමට එරෙහිව, සෙමින් අනුවර්තනය වන නිත්ය විසර්ජනය ලම්බක අඩු බලයක් පවත්වා ගෙන යන යාන්ත්රික උත්තේජනයක් මගින් හෝ වඩාත් ඵලදායී ලෙස චර්ම දිගු කිරීම මගින් ලබා ගනී. SA II ප්රතිචාරය ආරම්භ වන්නේ නොපැහැදිලි මායිම් සහිත විශාල ග්රාහක ක්ෂේත්ර වලින්. Ruffini receptors සමේ දිග හැරීමේ රටාව හරහා වස්තුවේ චලනයේ දිශාව අවබෝධ කර ගැනීමට දායක වේ [රූපය. 2 (A2)].
මීයන් තුළ, SA I සහ SA II ප්රතිචාර විද්යුත් භෞත විද්යාත්මකව වෙන් කළ හැක, ex-vivo ස්නායු සම සකස් කිරීමේදී. යාන්ත්රික සම්ප්රේෂණය සඳහා අවශ්ය දන්ත ඔස්මොටික් ශේෂය පවත්වා ගැනීම.
මීස්නර් කෝපුස්කල්. Meissner corpuscles ප්රධාන වශයෙන් අත්ල සහ පාද යටි පතුල්වල පමණක් නොව තොල්වල, දිවේ, මුහුණේ, තන පුඩුවල සහ ලිංගික අවයවවල ද ග්ලැසියර සමේ සමේ පැපිලා තුළ ස්ථානගත වී ඇත. ව්යුහ විද්යාත්මකව, ඒවා සංවෘත ස්නායු අවසානයකින් සමන්විත වේ, කැප්සියුලය සෑදී ඇත්තේ සම්බන්ධක පටක වල තැන්පත් කර ඇති තිරස් ලැමිලේ ලෙස සකස් කර ඇති පැතලි ආධාරක සෛල වලින් ය. තනි ස්නායු තන්තු A එකක් තිබේද? ශරීරයකට සම්බන්ධ වූ අනුකාරක [පය. 1 (C4)]. සිරුරේ ඕනෑම භෞතික විකෘතියක් ක්රියාකාරී විභවයන් විශාල ප්රමාණයක් අවුලුවන අතර එය ඉක්මනින් නතර වේ, එනම් ඒවා වේගයෙන් අනුවර්තනය වන ප්රතිග්රාහක වේ. උත්තේජකය ඉවත් කළ විට, ශරීරය නැවත එහි හැඩය ලබා ගන්නා අතර එසේ කරන අතරතුර තවත් ක්රියාකාරී විභවයන් ඇති කරයි. ඩර්මිස්හි ඒවායේ මතුපිට පිහිටීම හේතුවෙන්, මෙම සිරුරු සමේ චලනය, ස්ලිප් සහ කම්පන (20-40 Hz) ස්පර්ශයෙන් හඳුනා ගැනීම සඳහා තෝරා බේරා ප්රතිචාර දක්වයි. ඒවා ගතික සමට සංවේදී වේ - නිදසුනක් ලෙස, සම සහ හසුරුවන වස්තුවක් අතර [රූපය. 2 (A1)].
පැසීනියානු සිරුරු. පැසීනියානු කෝපස්කල් යනු සමේ ගැඹුරු යාන්ත්රික ප්රතිග්රාහක වන අතර සමේ චලිතයේ වඩාත් සංවේදී සංවෘත චර්ම යාන්ත්රික ප්රතිග්රාහක වේ. මෙම විශාල ඩිම්බ කෝෂ (දිග මි.මී. 1ක් දිග) තන්තුමය සම්බන්ධක පටකවල සංකේන්ද්රික ලැමිල්ලා සහ පැතලි වෙනස් කරන ලද ෂ්වාන් සෛල මගින් පෙලගැසී ඇති ෆයිබ්රොබ්ලාස්ට් ගැඹුරු ඩර්මිස් වල ප්රකාශ වේ. , එක A එකක් අවසන් කරයිද? afferent unmyelinated ස්නායු අවසානය [පය. 43 (C1)]. විශේෂයෙන් සංවේදී මධ්යස්ථානයක් සහිත සම මතුපිට විශාල ප්රතිග්රාහක ක්ෂේත්රයක් ඔවුන්ට ඇත. සී-මාෆ් විකෘති මීයන් තුළ වේගයෙන් අනුවර්තනය වන යාන්ත්රික ප්රතිග්රාහක වර්ග කිහිපයක සංවර්ධනය හා ක්රියාකාරිත්වය කඩාකප්පල් වේ. විශේෂයෙන්ම පැසීනියානු සිරුරු දැඩි ලෙස ක්ෂය වී ඇත.5
පැසීනියානු කෝපස්කල් සමේ ඉන්ඩෙන්ටේෂන් වලට ප්රතිචාර වශයෙන් ඉතා වේගවත් අනුවර්තනයක් පෙන්නුම් කරයි, වේගයෙන් අනුවර්තනය වන II (RA II) ස්නායු විසර්ජනය ඉහළ සංඛ්යාත කම්පන උත්තේජක අනුගමනය කිරීමට සමත් වන අතර සම්ප්රේෂණය වන කම්පන හරහා දුරස්ථ සිදුවීම් අවබෝධ කර ගැනීමට ඉඩ සලසයි. උත්තේජකයේ ආරම්භයේ සහ ඕෆ්සෙට් වලදී අස්ථිර ක්රියාකාරකම් සමඟ තිරසාර ඉන්ඩෙන්ටේෂන් වලට ප්රතිචාර දක්වයි. උත්තේජකයේ ප්රබලතාවයේ වෙනස්කම් හඳුනාගත හැකි නිසාත්, උත්තේජකයේ වෙනස් වීමේ වේගය වෙනස් කළහොත් (කම්පන වලදී සිදු වන පරිදි) ඒවායේ ප්රතිචාරය මෙම වෙනසට සමානුපාතික වන නිසාත් ඒවා ත්වරණ අනාවරක ලෙස හැඳින්වේ. Pacinian corpuscles දළ පීඩන වෙනස්වීම් සහ බොහෝ විට කම්පන (45-150 Hz) දැනෙනවා, ඒවා සෙන්ටිමීටර පවා ඈතින් හඳුනාගත හැකිය [රූපය. 300 (A2)].
ටොනික් ප්රතිචාරය decapsulated Pacinian corpuscle තුළ නිරීක්ෂණය විය.46 ඊට අමතරව, GABA-මැදිහත් වූ සංඥාව lamellate glia සහ ස්නායුවක් අතර අවහිර වූ විට, යාන්ත්රික සීමාවන් හෝ ප්රතිචාර සංඛ්යාතය වෙනස් නොකර, නියත ඉන්ඩෙන්ටේෂන් උත්තේජක වලදී නොවෙනස්ව පවතින Pacinian කෝපස්කල් තිරසාර ක්රියාකාරකම් සමඟ ප්රතිචාර දක්වයි. පැසීනියානු කෝපස්කල්හි නියුරෝන නොවන සංරචක යාන්ත්රික උත්තේජකය පෙරීමේදී මෙන්ම සංවේදක නියුරෝනවල ප්රතිචාර ගුණ වෙනස් කිරීමේදී ද්විත්ව භූමිකාවන් තිබිය හැකිය.
කොඳු ඇට පෙළේ ප්රක්ෂේපණ. සුෂුම්නාව තුළ ඇති A?-LTMR වල ප්රක්ෂේපන ශාඛා දෙකකට බෙදා ඇත. ප්රධාන මධ්යම ශාඛාව කොඳු ඇට පෙළේ ඉප්සිලේටරල් පෘෂ්ඨීය තීරු වල ගැබ්ගෙල මට්ටම දක්වා ඉහළ යයි [රූපය. 1 (B3)]. ද්විතියික ශාඛා ලැමිනේ IV හි පෘෂ්ඨීය අං වලින් අවසන් වන අතර වේදනාව සම්ප්රේෂණයට බාධා කරයි. මෙය ගේට්ටු පාලනයේ කොටසක් ලෙස වේදනාව අඩු කරයි [රූපය. 1 (B4)].48
ගැබ්ගෙල මට්ටම්වලදී, ප්රධාන ශාඛාවේ අක්සෝන පත්රිකා දෙකකින් වෙන් වේ: මධ්ය රේඛා පත්රිකාව ශරීරයේ පහළ භාගයෙන් (කකුල් සහ කඳ) තොරතුරු සම්ප්රේෂණය කරන ග්රැසිල් ෆැසිකල් වලින් සමන්විත වන අතර පිටත පත්රිකාව ඉහළ භාගයේ සිට තොරතුරු සම්ප්රේෂණය කරන උල් ෆැසිකල් වලින් සමන්විත වේ. ශරීරයේ (අත් සහ කඳ) [පය. 1 (B5)].
ප්රාථමික ස්පර්ශක අනුග්රහකයින් ඔවුන්ගේ පළමු උපාගමය සිදු කරන්නේ මෙඩුල්ලා හි දෙවන පෙළ නියුරෝන සමඟින් වන අතර එහිදී එක් එක් පත්රිකාවේ තන්තු එකම නමකින් යුත් න්යෂ්ටියක උපාගමනය කරයි: ග්රැසිල් න්යෂ්ටියේ ග්රැසිල් ෆැසිකුලස් ඇක්සෝන්ස් උපාගම සහ කියුනේට් ඇක්සෝන්ස් උපාගමය [යුක්ලියස් ඇක්සෝන්ස් උපාගමය. 1 (B6)]. උපාගමය ලබා ගන්නා නියුරෝන ද්විතියික අනුකාරක ලබා දෙන අතර මධ්ය රේඛාව තරණය කර මොළයේ කඳේ ප්රතිවිරුද්ධ පැත්තේ පත්රිකාවක් සාදයි - මධ්ය ලෙම්නිස්කස්, එය මොළයේ කඳ හරහා මැද මොළයේ ඊළඟ රිලේ ස්ථානයට, විශේෂයෙන්, තලමස් හි ඉහළ යයි. . 1 (B7)].
LTMR වල අණුක පිරිවිතර. LTMR වල මුල් විවිධාංගීකරණය පාලනය කරන අණුක යාන්ත්රණ මෑතකදී අර්ධ වශයෙන් පැහැදිලි කර ඇත. E2–11 කළල මීයන් DRG හි Ret tyrosine kinase receptor (Ret) සහ එහි සම ප්රතිග්රාහක GFR?13 ප්රකාශ කරන නියුරෝන ජනගහන මගින් පිටපත් කිරීමේ සාධකය Mafa.49,50 වරණාත්මකව සමීකරණය කරන බව Bourane සහ සහකාරකයින් පෙන්වා දී ඇත. Ret/GFR?2 නියුරෝන උපතේදී නිශ්චිත LTRM වර්ග තුනක් බවට පත්වීමට නියමිතයි: SA1 නියුරෝන මර්කල්-සෛල සංකීර්ණ නවීකරණය කිරීම, වේගයෙන් අනුවර්තනය වන නියුරෝන Meissner corpuscles නවීකරණය කිරීම සහ වේගයෙන් අනුවර්තනය වන afferents (RA I) රෝම කූප වටා ලාන්සිලේට් අවසානය සාදයි. Ginty සහ සහකාරකයන් ද වාර්තා කරන්නේ මුල්-රෙට් ප්රකාශ කරන DRG නියුරෝන Meissner corpuscles, Pacinian corpuscles සහ හිසකෙස් වටා ඇති ලැන්සිලේට් අන්තයන්ගෙන් යාන්ත්රික ප්රතිග්රාහක වේගයෙන් අනුවර්තනය වන බවයි. මොළයේ කඳේ නියුරෝන අක්ෂීය ප්රක්ෂේපණ.
මිනිස් සමේ යාන්ත්රික ප්රතිග්රාහක ගවේෂණය. 1968 දී Hagbarth සහ Vallbo විසින් විස්තර කරන ලද "Microneurography" තාක්ෂණය මාංශ පේශි, සන්ධි සහ සමට සපයන තනි මානව යාන්ත්රික සංවේදී අවසානයන්හි විසර්ජන හැසිරීම අධ්යයනය කිරීම සඳහා යොදා ගෙන ඇත (සමාලෝචනය සඳහා Macefield, 2005 බලන්න).52,53 මානව සමේ ක්ෂුද්ර නියුරෝග්රැෆි බහුතරයක්. අධ්යයනයන් මගින් අතේ ග්ලැසියර සමෙහි ඇති ස්පර්ශක ඇෆෙරන්ට් වල කායික විද්යාව සංලක්ෂිත කර ඇත. මිනිස් විෂයයන්හි මධ්ය සහ උල්නර් ස්නායු වලින් ක්ෂුද්ර ඉලෙක්ට්රෝඩ පටිගත කිරීම් LTMR වර්ග හතර විසින් ජනනය කරන ලද ස්පර්ශ සංවේදනය හෙළිදරව් කර ඇත: Meissner afferents සම හරහා ආලෝකයට පහර දීම, දේශීය කැපුම් බලවේගවලට ප්රතිචාර දැක්වීම සහ ප්රතිග්රාහක ක්ෂේත්රය තුළ ආරම්භක හෝ විවෘත ලිස්සා යාම සඳහා විශේෂයෙන් සංවේදී වේ. Pacinian afferents කඩිසර යාන්ත්රික සංක්රාන්ති වලට අතිශයින් සංවේදී වේ. ප්රතිග්රාහක ක්ෂේත්රය හරහා පිඹීමට ප්රබල ලෙස ප්රතිචාර දක්වයි. සංඛ්යාංකයක පිහිටා ඇති පැසීනියානු සිරුර සාමාන්යයෙන් අත ආධාරක මේසයට තට්ටු කිරීමට ප්රතිචාර දක්වයි. Merkel afferents ලාක්ෂණිකව විවික්ත ප්රදේශයකට යොදන ඉන්ඩෙන්ටේෂන් උත්තේජක සඳහා ඉහළ ගතික සංවේදීතාවයක් ඇති අතර බොහෝ විට මුදා හැරීමේදී අක්රිය විසර්ජනයක් සමඟ ප්රතිචාර දක්වයි. Ruffini afferents සාමාන්යයෙන් සමට යොදන බලවේග වලට ප්රතිචාර දක්වන නමුත්, SA II afferents හි සුවිශේෂී ලක්ෂණය වන්නේ පාර්ශ්වීය සමේ දිගු කිරීමට ප්රතිචාර දැක්වීමේ හැකියාවයි. අවසාන වශයෙන්, නළලෙහි හිසකෙස් ඒකක තනි හිසකෙස් වලට අනුරූප වන බහු සංවේදී ලප වලින් සමන්විත විශාල ඩිම්බකෝෂ හෝ අක්රමවත් ප්රතිග්රාහක ක්ෂේත්ර ඇත (එක් එක් afferent සැපයුම ~20 හිසකෙස්).
Keratinocytes හි යාන්ත්රික සංවේදීතාව
සම මත ඇති ඕනෑම යාන්ත්රික උත්තේජකයක් එපීඩර්මිස් සාදන keratinocytes හරහා සම්ප්රේෂණය කළ යුතුය. මෙම සර්වසම්පූර්ණ සෛල ඔවුන්ගේ ආධාරක හෝ ආරක්ෂිත භූමිකාවන්ට අමතරව සංඥා කාර්යයන් ඉටු කළ හැකිය. උදාහරණයක් ලෙස, keratinocytes යාන්ත්රික සහ ඔස්මොටික් උත්තේජක වලට ප්රතිචාර වශයෙන් වැදගත් සංවේදී සංඥා අණුවක් වන ATP ස්රාවය කරයි. -kinase signaling pathway සහ පසුව ඇති වූ F-actin ආතති තන්තු සෑදීම මගින් කෙරටිනොසයිට් වල යාන්ත්රික විරූපණය මර්කෙල් සෛල වැනි අසල්වැසි සෛල වලට යාන්ත්රිකව බාධා කළ හැකි බව යෝජනා කරයි හානිකර ස්පර්ශය සඳහා C-තන්තු රහිත අවසානය [Fig. 54,55 (C55)].1
හානිකර ස්පර්ශය
High threshold mechanoreceptors (HTMRs) යනු epidermal C- සහ A? නිදහස් ස්නායු අවසානය. ඒවා විශේෂිත ව්යුහයන් සමඟ සම්බන්ධ නොවන අතර හිසකෙස් සම යන දෙකෙහිම නිරීක්ෂණය කෙරේ [රූපය. 1 (A9)] සහ දිලිසෙන සම [රූපය. 1(C7)]. කෙසේ වෙතත්, ස්නායු අවසානය සෑම විටම keratinocyte හෝ Langherans සෛල හෝ melanocytes සමඟ සමීපව පවතින බැවින් නිදහස් ස්නායු අවසානය පිළිබඳ පදය විචක්ෂණශීලීව සලකා බැලිය යුතුය. ස්නායු අවසානය පිළිබඳ අල්ට්රාව්යුහාත්මක විශ්ලේෂණය මඟින් රළු එන්ඩොප්ලාස්මික් රෙටිකුලම්, බහුල මයිටොකොන්ඩ්රියා සහ ඝන-හරය වෙසිලිය ඇති බව හෙළි වේ. එපීඩර්මල් සෛලවල යාබද පටල ඝන වී ඇති අතර ස්නායු පටක වල පශ්චාත් උපාගමික පටලයට සමාන වේ. එපීඩර්මල් සෛල ATP, interleukine (IL6, IL10) සහ bradykinin ලෙස මැදිහත්කරුවන් මුදා හැරිය හැකි අතර අනෙක් අතට පෙප්ටයිඩර්ජික් ස්නායු අවසානයන් CGRP වැනි පෙප්ටයිඩ හෝ අපිචර්ම සෛල මත ක්රියා කරන P වැනි ද්රව්ය මුදා හැරිය හැකි බැවින් ස්නායු අවසානය සහ එපීඩර්මල් සෛල අතර අන්තර්ක්රියා ද්විපාර්ශ්වික විය හැකි බව සලකන්න. HTMRs සමන්විත වන්නේ හානිකර යාන්ත්රික උත්තේජක සහ පොලිමොඩල් nociceptors මගින් පමණක් උද්යෝගිමත් වන යාන්ත්රික-නොසිසෙප්ටර වලින් වන අතර ඒවා හානිකර තාපයට සහ බාහිර රසායනිකයට ප්රතිචාර දක්වයි. 2 (B2)].58
HTMR afferent තන්තු අවසන් වන්නේ සුෂුම්නාවෙහි පෘෂ්ඨීය අං තුළ ඇති ප්රක්ෂේපන නියුරෝන මතය. A?-HTMRs ප්රධාන වශයෙන් ලැමිනා I සහ V හි දෙවන පෙළ නියුරෝන සම්බන්ධ කරන අතර C-HTMR ලැමිනා II හි අවසන් වේ [රූපය. 1 (B8)]. දෙවන අනුපිළිවෙල nociceptive නියුරෝන සුෂුම්නාවේ පාලන පැත්තට ප්රක්ෂේපණය කර සුදු පදාර්ථයෙන් නැඟී anterolateral පද්ධතිය සාදයි. මෙම නියුරෝන ප්රධාන වශයෙන් තලමස් වල අවසන් වේ [රූපය. 1 (B9 සහ B10)].
Somatosensory නියුරෝන වල Mechano-ධාරා
යාන්ත්රික ප්රතිග්රාහක මන්දගාමී හෝ වේගවත් අනුවර්තනය වීමේ යාන්ත්රණයන් තවමත් පැහැදිලි කර නොමැත. සංවේදක ස්නායු අවසානයෙහි සෛලීය පරිසරය, යාන්ත්රිකව දොරටු සහිත නාලිකා වල ආවේණික ගුණාංග සහ සංවේදක නියුරෝනවල ඇති අක්ෂීය වෝල්ටීයතා-ගෙටඩ් අයන නාලිකා වල ගුණාංග මගින් යාන්ත්රික ප්රතිග්රාහක අනුවර්තනය කෙතරම් දුරට සපයන්නේද යන්න පැහැදිලි නැත (රූපය 2). කෙසේ වෙතත්, යාන්ත්රිකව දොරටු ධාරා වල ගුනාංගීකරනයේ මෑත කාලීන ප්රගතිය පෙන්නුම් කර ඇත්තේ DRG නියුරෝන වල විවිධ වර්ගයේ යාන්ත්රික සංවේදී නාලිකා පවතින බවත් යාන්ත්රික ප්රතිග්රාහක අනුවර්තනය වීමේ සමහර පැති පැහැදිලි කළ හැකි බවත්ය.
මීයන් තුළ වීට්රෝ පටිගත කිරීම මගින් පෙන්නුම් කෙරෙන්නේ DRG නියුරෝනවල සෝමා අභ්යන්තරව යාන්ත්රික සංවේදී සහ ප්රකාශිත කැටායන යාන්ත්රික ද්වාර ධාරා වන බවයි. සහ බෙන්සාමිල්, අර්ධ අවහිර කිරීමට හේතු වේ.59 FM64-60,62,63 කල් පවතින අවහිරකයක් ලෙස ක්රියා කරයි, සහ FM1-43 මීයන්ගේ පසුපස පාදයට එන්නත් කිරීම Randall-Selitto පරීක්ෂණයේදී වේදනා සංවේදීතාව අඩු කරන අතර තක්සේරු කරන ලද පාද ඉවත් කිරීමේ සීමාව වැඩි කරයි. von Frey හිසකෙස් සමග.1
තිරසාර යාන්ත්රික උත්තේජනයට ප්රතිචාර වශයෙන්, වසා දැමීම හරහා යාන්ත්රික සංවේදී ධාරා අඩු වේ. ධාරා ක්ෂය වීමේ කාල නියතයන් මත පදනම්ව, යාන්ත්රික සංවේදී ධාරා වර්ග හතරක් වෙන්කර හඳුනාගත හැකිය: වේගයෙන් අනුවර්තනය වන ධාරා (~ 3~6 ms), අතරමැදි ලෙස අනුවර්තනය වන ධාරා (~15-30 ms), සෙමින් අනුවර්තනය වන ධාරා (~200-300 ms ) සහ අතිශය සෙමින් අනුවර්තනය වන ධාරා (~ 1000 ms).64 මෙම සියලුම ධාරා මීයන් DRG නියුරෝන වල විචල්ය සිදුවීම් සමඟින් ඇති අතර එය පසුපස පාදයේ ග්ලැසියර සම නවීකරණය කරයි.
සාපේක්ෂ සවිස්තරාත්මක උත්තේජක-ධාරා විශ්ලේෂණයට ඉඩ සලසන වර්ධක යාන්ත්රික උත්තේජක මාලාවක් යෙදීමෙන් යාන්ත්රික සංවේදී ධාරා වල යාන්ත්රික සංවේදිතාව තීරණය කළ හැක. එකවර විවෘතව පවතින නාලිකා.66 සිත්ගන්නා කරුණ නම්, ඉතා සෙමින් අනුවර්තනය වන යාන්ත්රික සංවේදී ධාරාව හා සසඳන විට වේගයෙන් අනුවර්තනය වන යාන්ත්රික සංවේදී ධාරාව අඩු යාන්ත්රික එළිපත්ත සහ අර්ධ-සක්රීය මධ්ය ලක්ෂ්යය දර්ශණය වන බව වාර්තා වී ඇත.64,67
Nociceptive phenotypes සහිත සංවේදී නියුරෝන අඩු යාන්ත්රික සීමාවක් සහිත ශීඝ්රයෙන් අනුවර්තනය වන යාන්ත්රික සංවේදී ධාරා ප්රකාශ කරයි. මෙම ධාරා vivo හි LTMR සහ HTMR වල දක්නට ලැබෙන විවිධ යාන්ත්රික සීමාවන්ට දායක විය හැකි බවට මෙය යෝජනා කළේය. මෙම in vitro අත්හදා බැලීම් ප්රවේශමෙන් කළ යුතු වුවද, අඩු සහ ඉහළ සීමාවන් ඇති යාන්ත්රික සම්ප්රේෂකවල DRG නියුරෝන වල සෝමා වල පැවැත්ම සඳහා සහය සපයනු ලැබුවේ සංස්කෘතික මූසික සංවේදී නියුරෝනවල රේඩියල් ස්ට්රෙච්-පාදක උත්තේජනය මගිනි.60,61,63,64,68 මෙම සුසමාදර්ශය දෙකක් අනාවරණය කළේය. දිගු-සංවේදී නියුරෝන වල ප්රධාන ජනගහන, එකක් අඩු උත්තේජක විස්තාරයට ප්රතිචාර දක්වන අතර තවත් එකක් ඉහළ උත්තේජක විස්තාරයට තෝරා ප්රතිචාර දක්වයි.
මෙම ප්රතිඵලවලට වැදගත්, නමුත් සමපේක්ෂන, යාන්ත්රික ඇඟවුම් ඇත: සංවේදක නියුරෝනවල යාන්ත්රික එළිපත්ත යාන්ත්රික ප්රතිග්රාහකයේ සෛලීය සංවිධානය සමඟ එතරම් සම්බන්ධයක් නොතිබිය හැකි නමුත් යාන්ත්රිකව දොරටු සහිත අයන නාලිකාවල ගුණ තුළ පවතී.
මී ඩීආර්ජී නියුරෝනවල යාන්ත්රික සංවේදී කැටායන ධාරා වල සංවේදිකරණයට පාදක වන යාන්ත්රණයන් මෑතකදී අනාවරණය කර ඇත.64,67 එය නාලිකා ගුණාංගවලට බලපාන සමගාමී යාන්ත්රණ දෙකකින් ප්රතිඵල ලබයි: අනුවර්තනය සහ අක්රිය වීම. අනුවර්තනය වීම මුලින්ම වාර්තා වූයේ ශ්රවණ කෙස් සෛල අධ්යයනයේදීය. එය යාන්ත්රික උත්තේජක අක්ෂය ඔස්සේ පරිවර්තක නාලිකාවේ සක්රීය වක්රයේ සරල පරිවර්තනයක් ලෙස ක්රියාකාරීව විස්තර කළ හැක.70-72 අනුවර්තනය මඟින් සංවේදී ප්රතිග්රාහකයන්ට පවතින උත්තේජකයක් ඉදිරියේ නව උත්තේජකවලට ඔවුන්ගේ සංවේදීතාව පවත්වා ගැනීමට ඉඩ සලසයි. කෙසේ වෙතත්, DRG නියුරෝනවල යාන්ත්රික සංවේදී ධාරා වල සැලකිය යුතු කොටසක් සමීකරණ යාන්ත්රික උත්තේජනයෙන් පසුව නැවත සක්රිය කළ නොහැක, සමහර පරිවර්තක නාලිකා අක්රිය වීම පෙන්නුම් කරයි. මෙම යාන්ත්රණයන් දෙක මී ඩීආර්ජී නියුරෝන තුළ හඳුනාගෙන ඇති සියලුම යාන්ත්රික සංවේදී ධාරා සඳහා පොදු වන අතර, අදාළ භෞතික රසායනික මූලද්රව්ය මෙම නාලිකාවල චාලකත්වය තීරණය කරන බව යෝජනා කරයි.64,67
අවසාන වශයෙන්, අණුක මට්ටමින් සම්ප්රේෂණ යාන්ත්රණයන් හඳුනා ගැනීමේ ගවේෂණයේ දී vitro හි ආවේණික යාන්ත්රික සංවේදී ධාරා වල ගුණ නිර්ණය කිරීම ඉතා වැදගත් වේ. ඩීආර්ජී නියුරෝනවල යාන්ත්රික එළිපත්තෙහි සහ අනුවර්තනය වන චාලක විද්යාවේ නිරීක්ෂණය කරන ලද විචල්යතාවයෙන් ඇඟවෙන්නේ අයන නාලිකාවල ආවේණික ගුණාංග අවම වශයෙන් අර්ධ වශයෙන් යාන්ත්රික එළිපත්ත සහ 1960 දශකවල විස්තර කරන ලද යාන්ත්රික ප්රතිග්රාහකවල අනුවර්තන චාලක විද්යාව පැහැදිලි කළ හැකි බවයි. 80 ex vivo සූදානම භාවිතා කරයි.
යාන්ත්රික සංවේදී ප්රෝටීන
Somatosensory නියුරෝන වල යාන්ත්රික සංවේදී අයන ධාරා හොඳින් සංලක්ෂිත වේ, ඊට ප්රතිවිරුද්ධව, ක්ෂීරපායින් තුළ යාන්ත්රික සම්ප්රේෂණයට මැදිහත් වන අණු වල අනන්යතාවය ගැන එතරම් දැනුමක් නැත. Drosophila සහ C. elegans හි ජාන තිර මගින් TRP සහ degenerin/epithelial Na+ channel (Deg/ENaC) පවුල් ඇතුළුව අපේක්ෂක යාන්ත්රික සම්ප්රේෂණ අණු හඳුනාගෙන ඇත. . මීට අමතරව, මෙම අපේක්ෂකයින් බොහෝමයක් චර්ම යාන්ත්රික ප්රතිග්රාහක සහ සෝමැටෝසෙන්සරි නියුරෝන වල පවතී (රූපය 73).
අම්ල සංවේදී අයන නාලිකා
ASICs degenerin-epithelial Na+ නාලිකා පවුලේ ප්රෝටෝන දොරටු සහිත උප සමූහයකට අයත් වේ.74 ASIC පවුලේ සාමාජිකයින් තිදෙනෙකු (ASIC1, ASIC2 සහ ASIC3) යාන්ත්රික ප්රතිග්රාහක සහ nociceptors වලින් ප්රකාශ වේ. ASIC නාලිකා වල කාර්යභාරය ASIC නාලිකා ජානවල ඉලක්කගත මකාදැමීම් සහිත මීයන් භාවිතා කරමින් චර්යාත්මක අධ්යයනයන්හි විමර්ශනය කර ඇත. ASIC1 මකා දැමීම චර්ම යාන්ත්රික ප්රතිග්රාහකවල ක්රියාකාරිත්වය වෙනස් නොකරන නමුත් අන්ත්රය නවීකරණය කරන අනුකාරකයන්ගේ යාන්ත්රික සංවේදීතාව වැඩි කරයි. visceral mechano-nociception සහ cutaneous mechanosensation යන දෙකම.75 ASIC2 කඩාකප්පල් කිරීම visceral afferents හි යාන්ත්රික සංවේදීතාව අඩු කරන අතර චර්ම HTMRs හානිකර උත්තේජක වලට ප්රතිචාර දැක්වීම අඩු කරයි.76
සංක්රාන්ති ප්රතිග්රාහක නාලිකාව
THE TRP සුපිරි පවුල ක්ෂීරපායීන්ගේ උප පවුල් හයකට බෙදා ඇත.78 සියලුම TRP උපපවුල්වල විවිධ සෛල පද්ධතිවල යාන්ත්රික සංවේදනයට සම්බන්ධ වූ සාමාජිකයින් ඇත. සහ TRP නාලිකා දෙකක් පමණක්, TRPV79 සහ TRPA4, ස්පර්ශ ප්රතිචාර දැක්වීමට සම්බන්ධ වී ඇත. මීයන් තුළ TRPV1 ප්රකාශනය කඩාකප්පල් කිරීම උග්ර යාන්ත්රික සංවේදන සීමාවන් කෙරෙහි සාමාන්ය බලපෑමක් ඇති කරයි, නමුත් හානිකර යාන්ත්රික උත්තේජක සඳහා දැඩි ලෙස සංවේදීතාව අඩු කරයි. 4 TRPA80,81 යාන්ත්රික හයිපර්ල්ජීසියාවේ භූමිකාවක් ඇති බව පෙනේ. TRPA4-අඩු මීයන් වේදනා අධි සංවේදීතාව විදහා දක්වයි. TRPA82,83 nociceptor සංවේදක නියුරෝන තුළ යාන්ත්රික, සීතල සහ රසායනික උත්තේජක සම්ප්රේෂණය කිරීමට දායක වන නමුත් එය හිසකෙස්-සෛල සම්ප්රේෂණය සඳහා අත්යවශ්ය නොවන බව පෙනේ.1
ක්ෂීරපායින් තුළ ප්රකාශිත TRP නාලිකා සහ ASIC නාලිකා යාන්ත්රිකව දොරටු ඇති බවට පැහැදිලි සාක්ෂි නොමැත. මෙම නාලිකා කිසිවක් විෂමජාතීය ලෙස ප්රකාශිත ඒවායේ ස්වදේශික පරිසරයේ නිරීක්ෂණය කරන ලද යාන්ත්රික සංවේදී ධාරා වල විද්යුත් අත්සන නැවත සංග්රහ නොකරයි. යාන්ත්රික සම්ප්රේෂණ නාලිකාවක් එහි සෛලීය සන්දර්භයෙන් පිටත ක්රියා කළ හැකිද යන්න පිළිබඳ අවිනිශ්චිතතාවය සැලකිල්ලට ගෙන, ASICs සහ TRPs නාලිකා යාන්ත්රික සම්ප්රේෂකයන් වීමේ හැකියාව බැහැර නොකරයි (SLP3 කොටස බලන්න).
Piezo ප්රෝටීන්
Coste සහ සහකාරකයින් විසින් Mechanosensing ප්රෝටීන සඳහා පොරොන්දු වූ අපේක්ෂකයින් ලෙස Piezo protiens මෑතකදී හඳුනාගෙන ඇත. . Piezo 86,87 DRG වල බහුලව ඇති අතර Piezo 1 යන්තම් හඳුනාගත නොහැක. Piezo-ප්රේරිත යාන්ත්රික සංවේදී ධාරාවන් gadolinium, ruthenium red සහ GsMTx2 (ටැරන්ටුලා ග්රැම්මෝස්ටෝලා ස්පාටුලටා වෙතින් විෂ වර්ගයක්) මගින් වලක්වනු ලැබේ. Piezo 38 ට වඩා. අන්තරාසර්ග යාන්ත්රික සංවේදී ධාරා වලට සමානව, Piezo මත යැපෙන ධාරා වලට 38 mV පමණ ප්රතිවර්තන විභවයන් ඇති අතර Na+, K+, Ca2+ සහ Mg1+ යන සියල්ලම යටින් පවතින නාලිකාව හරහා විහිදෙන කැටායන තේරීමක් නොමැත. ඒ හා සමානව, piezo මත යැපෙන ධාරා පටල විභවය මගින් නියාමනය කරනු ලැබේ, depolarized විභවයන්හිදී ධාරා චාලකයේ කැපී පෙනෙන මන්දගාමී වීමකි.4
Piezo ප්රෝටීන නිසැකවම යාන්ත්රික සංවේදී ප්රෝටීන වන අතර සංවේදී නියුරෝනවල යාන්ත්රික සංවේදී ධාරා වේගයෙන් අනුවර්තනය වීමේ බොහෝ ගුණාංග බෙදා ගනී. Cultered DRG නියුරෝන සඳහා Piezo 2 short interfering RNA සමඟ ප්රතිකාර කිරීම වේගයෙන් අනුවර්තනය වන ධාරාවක් සහිත නියුරෝනවල අනුපාතය අඩු වූ අතර යාන්ත්රික සංවේදී නියුරෝන ප්රතිශතය අඩු විය. හඳුනාගෙන ඇත. කෙසේ වෙතත්, Mouse Piezo 86 ප්රෝටීනය පිරිසිදු කර අසමමිතික ලිපිඩ ද්වී ස්ථරවලට ප්රතිනිර්මාණය කර රුතේනියම් රතු වලට සංවේදී අයන නාලිකා liposome සාදයි. තනි Piezo සාමාජිකයා මකා දැමීමෙන් සාමාන්ය ස්පර්ශයට බල නොපා හානිකර උත්තේජකවලට යාන්ත්රික ප්රතිචාරය අඩු කළ ඩ්රොසෝෆිලා හි තොරතුරු ලබා දී ඇත. ස්පර්ශ සංවේදනය. උදාහරණයක් ලෙස, රක්තහීනතාවයෙන් පෙළෙන රෝගීන් (පරම්පරාගත xerocytosis) පිළිබඳ මෑත අධ්යයනයකින් පෙන්නුම් කරන්නේ එරිත්රෝසයිට් පරිමාව හෝමියස්ටැසිස් පවත්වා ගැනීම සඳහා Piezo 1 හි භූමිකාවයි.87
Transmembrane Channel-Like (TMC)
මෑත කාලීන අධ්යයනයකින් පෙන්නුම් කරන්නේ හිසකෙස් සෛල යාන්ත්රික සම්ප්රේෂණය සඳහා TMC1 සහ TMC2 යන ප්රෝටීන දෙකක් අවශ්ය වන බවයි.91 TMC1 ජාන විකෘතිය හේතුවෙන් පාරම්පරික බිහිරි භාවය මිනිසුන් සහ මීයන් තුළ වාර්තා විය. , නමුත් එය විමර්ශනය කිරීමට හොඳ මඟ පෙන්වීමක් බව පෙනේ.
ස්ටෝමැටින් වැනි ප්රෝටීන් 3 (SLP3)
සම්ප්රේෂණ නාලිකාවලට අමතරව, නාලිකාවට සම්බන්ධ සමහර අමතර ප්රෝටීන ස්පර්ශ සංවේදීතාවයේ කාර්යභාරයක් ඉටු කරන බව පෙන්වා දී ඇත. SLP3 ක්ෂීරපායී DRG නියුරෝන වල ප්රකාශ වේ. SLP3 නොමැති විකෘති මීයන් යොදාගෙන කරන ලද අධ්යයනයන් මගින් යාන්ත්රික සංවේදනය සහ යාන්ත්රික ධාරා වල වෙනසක් පෙන්නුම් කර ඇත.94,95 SLP3 නිරවද්ය ක්රියාකාරිත්වය තවමත් නොදනී. එය එහි C. elegans homolog MEC2.96 මෑතකදී GR සඳහා යෝජනා කර ඇති පරිදි යාන්ත්රික සංවේදී නාලිකාව සහ යටින් පවතින ක්ෂුද්ර නල අතර සම්බන්ධකයක් විය හැකිය. ලෙවින් විද්යාගාරය DRG සංවේදක නියුරෝන මගින් ටෙදරයක් සංස්ලේෂණය කර යාන්ත්රික සංවේදී අයන නාලිකාව බාහිර සෛල අනුකෘතියට සම්බන්ධ කරයි. RA-යාන්ත්රික සංවේදී ධාරා, keratinocytes මගින් නිපදවන න්යාස ප්රෝටීනයක් වන laminin-97 මගින් ද නිෂේධනය කරයි, බාහිර සෛලීය ප්රෝටීන මගින් යාන්ත්රික සංවේදී ධාරාවේ මොඩියුලේෂන් කල්පිතය ශක්තිමත් කරයි.332
K+ නාලිකා උප පවුල
කැටායන වි ධ්රැවීකරණ යාන්ත්රික සංවේදී ධාරා වලට සමාන්තරව, ප්රතිධ්රැවීකරණය කරන යාන්ත්රික සංවේදී K+ ධාරා පැවතීම විමර්ශනය වෙමින් පවතී. යාන්ත්රික සංවේදී සෛලවල K+ නාලිකා වත්මන් සමතුලිතතාවයට පියවර ගත හැකි අතර යාන්ත්රික එළිපත්ත සහ යාන්ත්රික ප්රතිග්රාහක අනුවර්තනය වීමේ කාල පරාසය නිර්වචනය කිරීමට දායක වේ.
KCNK සාමාජිකයින් අයත් වන්නේ සිදුරු දෙකේ වසම K+ නාලිකාව (K2P) පවුලටය.99,100 K2P pH වෙනස්වීම්, තාපය, දිගු කිරීම සහ පටල විරූපණය ඇතුළුව සෛලීය, භෞතික සහ ඖෂධීය කාරක මගින් කැපී පෙනෙන නියාමනය පරාසයක් පෙන්වයි. මෙම K2P විවේක පටල විභවය තුළ ක්රියාකාරී වේ. KCNK උප ඒකක කිහිපයක් Somatosensory නියුරෝන වල ප්රකාශ වේ.101 KCNK2 (TREK-1), KCNK4 (TRAAK) සහ TREK-2 නාලිකා පටල දිගුවකින් සෘජු යාන්ත්රික ද්වාරයක් පෙන්වා ඇති නාලිකා කිහිපය අතර වේ.102,103.
කඩාකප්පල් වූ KCNK2 ජානයක් සහිත මීයන් තාපය සහ මෘදු යාන්ත්රික උත්තේජක සඳහා වැඩි දියුණු කරන ලද සංවේදීතාවයක් පෙන්නුම් කරයි, නමුත් Randall-Selitto පරීක්ෂණය භාවිතයෙන් hindpaw වෙත යොදන ලද හානිකර යාන්ත්රික පීඩනය සඳහා සාමාන්ය ඉවත් වීමේ සීමාවක් පෙන්නුම් කරයි. කොන්දේසි. KCNK104 knockout මීයන් මෘදු යාන්ත්රික උත්තේජනයට අධි සංවේදී වූ අතර, KCNK2 අතිරේක අක්රිය කිරීම මගින් මෙම අධි සංවේදීතාව වැඩි කරන ලදී. වෝල්ටීයතා දොරටු ධාරා depolarizing සහ repolarizing.
KCNK18 (TRESK) යනු සෝමැටෝසෙන්සරි නියුරෝනවල විවේක පටල විභවය නියාමනය කරන පසුබිම් K+ සන්නායකතාවයට ප්රධාන දායකයෙකි. මෙන්ම වේදනාකාරී යාන්ත්රික උත්තේජක. KCNK106 සහ තරමක් දුරට KCNK18, හයිඩ්රොක්සි-?-සැන්ෂූල් හි අණුක ඉලක්කය වීමට යෝජිතය, එය ෂෙසුආන් ගම්මිරිස්වල ඇති සංයෝගයක් වන අතර එය ස්පර්ශ ප්රතිග්රාහක සක්රීය කරන අතර මිනිසුන් තුළ හිරි වැටීමක් ඇති කරයි.18.
වෝල්ටීයතා රඳා පවතින K+ නාලිකාව KCNQ4 (Kv7.4) මීයන් සහ මිනිසුන් යන දෙඅංශයේම වේගයෙන් අනුවර්තනය වන යාන්ත්රික ප්රතිග්රාහකවල උප ජනගහනයක ප්රවේගය සහ සංඛ්යාත මනාපය සැකසීම සඳහා තීරණාත්මක වේ. KCNQ4 හි විකෘතිය මුලදී පාරම්පරික බිහිරි භාවයක් සමඟ සම්බන්ධ වී ඇත. මෑත කාලීන අධ්යයනයකින් KCNQ4 ස්ථානගත කර ඇත්තේ චර්ම ශීඝ්රයෙන් අනුවර්තනය වන රෝම කූපයේ සහ Meissner corpuscle වල පර්යන්ත ස්නායු අවසානයයි. ඒ අනුව, KCNQ4 ශ්රිතය නැතිවීම අඩු සංඛ්යාත කම්පනය සඳහා යාන්ත්රික ප්රතිග්රාහක සංවේදීතාව වරණාත්මකව වැඩි දියුණු කිරීමට හේතු වේ. කැපී පෙනෙන ලෙස, KCNQ4 ජානයේ ප්රමුඛ විකෘති හේතුවෙන් ප්රමාද වූ ශ්රවණාබාධ ඇති පුද්ගලයින් කුඩා-විස්තාරය, අඩු-සංඛ්යාත කම්පනය හඳුනාගැනීමේදී වැඩි දියුණු කළ කාර්ය සාධනයක් පෙන්නුම් කරයි.109
ආචාර්ය ඇලෙක්ස් ජිමිනෙස්ගේ තීක්ෂ්ණ බුද්ධිය
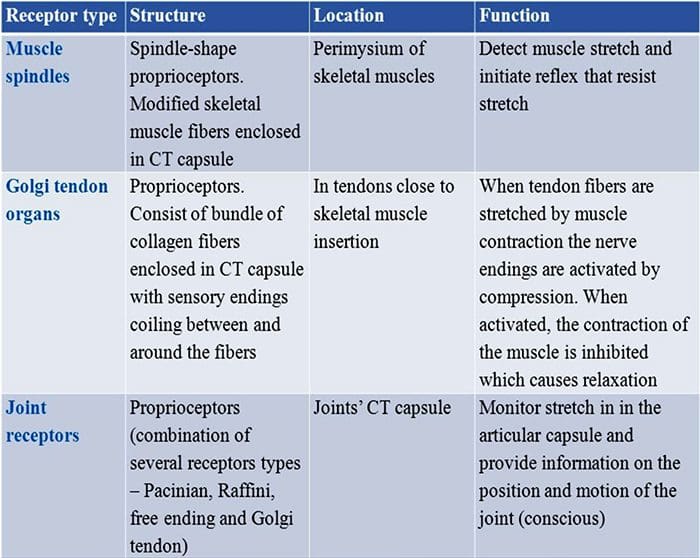
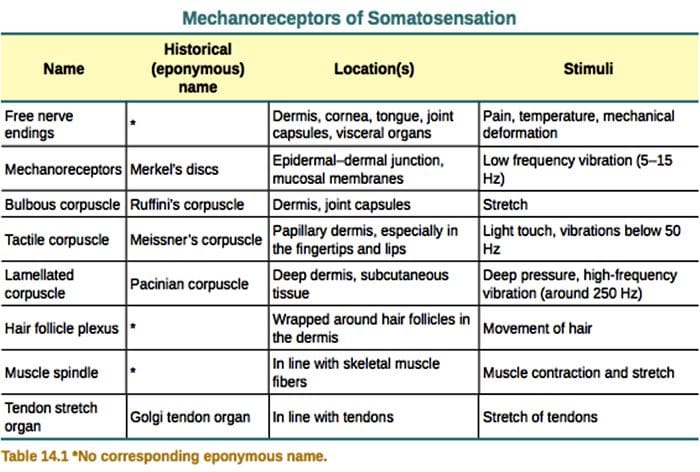
ස්පර්ශය මිනිස් සිරුරේ ඇති සංකීර්ණම ඉන්ද්රියයන්ගෙන් එකක් ලෙස සැලකේ, විශේෂයෙන්ම එය භාරව සිටින නිශ්චිත ඉන්ද්රියයක් නොමැති නිසා. ඒ වෙනුවට, ස්පර්ශ සංවේදනය සිදු වන්නේ සම හරහා ඇති යාන්ත්රික පීඩනයට හෝ විකෘතියට ප්රතිචාර දක්වන යාන්ත්රික ප්රතිග්රාහක ලෙස හඳුන්වන සංවේදක ප්රතිග්රාහක හරහා ය. ක්ෂීරපායීන්ගේ ග්ලැසියර හෝ හිසකෙස් නැති සමෙහි යාන්ත්රික ප්රතිග්රාහක ප්රධාන වර්ග හතරක් ඇත: ලැමිලර් කෝපස්කල්, ස්පර්ශක කෝපස්කල්, මර්කල් ස්නායු අවසානය සහ බල්බස් කෝපස්කල්. යාන්ත්රික ප්රතිග්රාහක ක්රියා කරන්නේ ස්පර්ශය හඳුනා ගැනීමට ඉඩ දීම සඳහා, ප්රොප්රියෝසෙප්ෂන් ලෙස හැඳින්වෙන මාංශ පේශි, අස්ථි සහ සන්ධිවල පිහිටීම නිරීක්ෂණය කිරීමට සහ ශරීරයේ ශබ්දය සහ චලනය පවා හඳුනා ගැනීමට ය. මෙම යාන්ත්රික ප්රතිග්රාහකවල ව්යුහයේ සහ ක්රියාකාරීත්වයේ යාන්ත්රණයන් අවබෝධ කර ගැනීම වේදනා කළමනාකරණය සඳහා ප්රතිකාර සහ ප්රතිකාර භාවිතා කිරීමේ මූලික අංගයකි.
නිගමනය
ස්පර්ශය යනු විවිධ වෙනස් කොට සැලකීමේ කාර්ය සාධනය සමඟ කම්පනය, හැඩය, වයනය, සතුට සහ වේදනාව යන විවිධ ස්පර්ශ ගුණාංග නියෝජනය කරන නිසා එය සංකීර්ණ හැඟීමකි. මේ දක්වා, ස්පර්ශ ඉන්ද්රියයක් සහ මනෝ භෞතික ඉන්ද්රිය අතර ලිපි හුවමාරුව සහසම්බන්ධ වූ අතර පන්ති විශේෂිත අණුක සලකුණු මතුවෙමින් තිබේ. අනාගත ප්රවේණි විද්යාව හඳුනාගැනීම පහසු කිරීම සඳහා ස්පර්ශ හැසිරීම් වල විවිධත්වයට ගැළපෙන මීයන් පරීක්ෂණ සංවර්ධනය කිරීම දැන් අවශ්ය වේ. සංවේදී afferent වර්ගවල නිශ්චිත උප කුලක නොමැති මීයන් භාවිතා කිරීම විශේෂිත ස්පර්ශක ක්රමයක් හා සම්බන්ධ යාන්ත්රික ප්රතිග්රාහක සහ සංවේදී තන්තු හඳුනා ගැනීමට බෙහෙවින් පහසුකම් සපයයි. සිත්ගන්නා කරුණ නම්, මෑත කාලීන පත්රිකාවක් මිනිසාගේ යාන්ත්රික සංවේදී ගතිලක්ෂණවල ප්රවේණික පදනම පිළිබඳ වැදගත් ප්රශ්නය විවෘත කරන අතර තනි ජාන විකෘතියක් ස්පර්ශ සංවේදීතාවයට අහිතකර ලෙස බලපෑ හැකි බව යෝජනා කරයි. ස්පර්ශ ක්රමයකට හෝ ස්පර්ශ හිඟයකට සම්බන්ධ සංවේදී නියුරෝනවල උප කුලකය නිශ්චිතව හඳුනා ගැනීමෙන් ප්රගතිය.
ඒ වෙනුවට, යාන්ත්රික දොරටු ධාරා වල ජෛව භෞතික ගුණාංග නිර්වචනය කිරීමට ප්රගතියක් ලබා ඇත. 64 මෑත වසරවල නව තාක්ෂණික ක්රම දියුණු කිරීම, පටල ආතති වෙනස්කම් නිරීක්ෂණය කිරීමට ඉඩ සලසයි, යාන්ත්රික-ගෙටඩ් ධාරාව පටිගත කරන අතරම, විස්තර කිරීමට වටිනා පර්යේෂණාත්මක ක්රමයක් ඔප්පු කර ඇත. වේගවත්, අතරමැදි සහ මන්දගාමී අනුවර්තනය සහිත යාන්ත්රික සංවේදී ධාරා (ඩෙල්මාස් සහ සහයෝගිතාකරුවන් තුළ සමාලෝචනය කර ඇත) 66,111 අනාගතයේදී ක්රියාකාරී විවිධ යාන්ත්රික ප්රතිග්රාහක අනුවර්තනය වීමේ යාන්ත්රණයන්හි වත්මන් ගුණාංගවල කාර්යභාරය සහ යාන්ත්රික සංවේදී K+ ධාරා වල උද්වේගකාරී බව තීරණය කරනු ඇත. LTMRs සහ HTMRs.
ක්ෂීරපායීන්ගේ යාන්ත්රික ධාරා වල අණුක ස්වභාවය ද අනාගත බලාපොරොත්තු සහගත පර්යේෂණ මාතෘකාවකි. අනාගත පර්යේෂණ ඉදිරිදර්ශන දෙකකින් ප්රගතියට පත් වනු ඇත, පළමුව සයිටොස්කෙලිටන් වෙත නාලිකා සම්බන්ධ කරන උපාංග අණුවේ කාර්යභාරය තීරණය කිරීමට සහ TRP සහ ASIC/EnaC පවුල් වැනි අයන නාලිකාවල යාන්ත්රික සංවේදීතාව ප්රදානය කිරීමට හෝ නියාමනය කිරීමට අවශ්ය වනු ඇත. දෙවනුව, ප්රධාන ප්රශ්නවලට පිළිතුරු සැපයීමෙන් Piezo නාලිකාවල දායකත්වයේ විශාල සහ පොරොන්දු වූ ක්ෂේත්රය විමර්ශනය කිරීම, විනිවිද යාම සහ ද්වාර යාන්ත්රණයන්, සංවේදක නියුරෝනවල උප කුලකය සහ Piezo සම්බන්ධ ස්පර්ශ ක්රම සහ නියුරෝන නොවන සෛල තුළ Piezo භූමිකාව යාන්ත්රික සංවේදනය.
මෙම සංවේදනයන් සැකසීමට නිශ්චිත ඉන්ද්රියයන් භාවිතා කරන දර්ශනය, රසය, ශබ්දය සහ සුවඳ හා සසඳන විට ස්පර්ශය පිළිබඳ හැඟීම, යාන්ත්රික ප්රතිග්රාහක ලෙස හඳුන්වන කුඩා ප්රතිග්රාහක හරහා ශරීරය පුරා ඇති විය හැක. විවිධ වර්ගයේ යාන්ත්රික ප්රතිග්රාහක සමෙහි විවිධ ස්ථරවල දක්නට ලැබෙන අතර එහිදී ඒවාට යාන්ත්රික උත්තේජක පුළුල් පරාසයක් හඳුනාගත හැකිය. ඉහත ලිපිය මගින් ස්පර්ශ සංවේදනය හා සම්බන්ධ යාන්ත්රික ප්රතිග්රාහකවල ව්යුහාත්මක සහ ක්රියාකාරී යාන්ත්රණවල ප්රගතිය පෙන්නුම් කරන විශේෂිත උද්දීපනයන් විස්තර කරයි. ජෛව තාක්ෂණ තොරතුරු ජාතික මධ්යස්ථානයෙන් (NCBI) තොරතුරු යොමු කර ඇත. අපගේ තොරතුරු වල විෂය පථය චිරොක්ට්රැක්ටික් මෙන්ම කොඳු ඇට පෙළේ තුවාල සහ තත්වයන්ට සීමා වේ. විෂය කරුණු සාකච්ඡා කිරීමට, කරුණාකර වෛද්ය ජිමිනෙස්ගෙන් විමසීමට හෝ අපව අමතන්න915-850-0900 .
ආචාර්ය ඇලෙක්ස් ජිම්ීනෙස් විසින් පාලනය කරන ලදී
අමතර මාතෘකා: පිටුපස වේදනාව
ආපහු වේදනාව ආබාධිතභාවයට සහ ලොව පුරා වැඩ කරන දින මග හැරීමට වඩාත් ප්රචලිත හේතුවකි. ඇත්ත වශයෙන්ම, වෛද්ය කාර්යාල චාරිකා සඳහා දෙවන වඩාත් පොදු හේතුව ලෙස කොන්දේ වේදනාව ආරෝපණය කර ඇති අතර එය ඉහළ ශ්වසන ආසාදනවලින් පමණක් වැඩි වේ. ජනගහනයෙන් ආසන්න වශයෙන් සියයට 80 කට පමණ ඔවුන්ගේ ජීවිත කාලය පුරාම අවම වශයෙන් එක් වරක්වත් යම් ආකාරයක පිටුපස වේදනාවක් අත්විඳිනු ඇත. කොඳු ඇට පෙළ යනු අනෙකුත් මෘදු පටක අතර අස්ථි, සන්ධි, බන්ධන සහ මාංශ පේශි වලින් සමන්විත සංකීර්ණ ව්යුහයකි. මේ නිසා, තුවාල සහ/හෝ උග්ර තත්ත්වයන්, වැනි හර්නිටීස් තැටි , අවසානයේ පිටුපස වේදනාවේ රෝග ලක්ෂණ ඇති විය හැක. ක්රීඩා තුවාල හෝ මෝටර් රථ අනතුරු තුවාල බොහෝ විට පිටුපස වේදනාව සඳහා නිතර නිතර හේතුව වේ, කෙසේ වෙතත්, සමහර විට සරලම චලනයන් වේදනාකාරී ප්රතිඵල ඇති විය හැක. වාසනාවකට මෙන්, චිරොක්ට්රැක්ටික් සත්කාර වැනි විකල්ප ප්රතිකාර විකල්ප, කොඳු ඇට පෙළ ගැලපීම් සහ අතින් හැසිරවීම් භාවිතයෙන් පිටුපස වේදනාව සමනය කිරීමට උපකාරී වන අතර අවසානයේ වේදනා සහන වැඩි දියුණු කරයි.
VIDEO
VIDEO

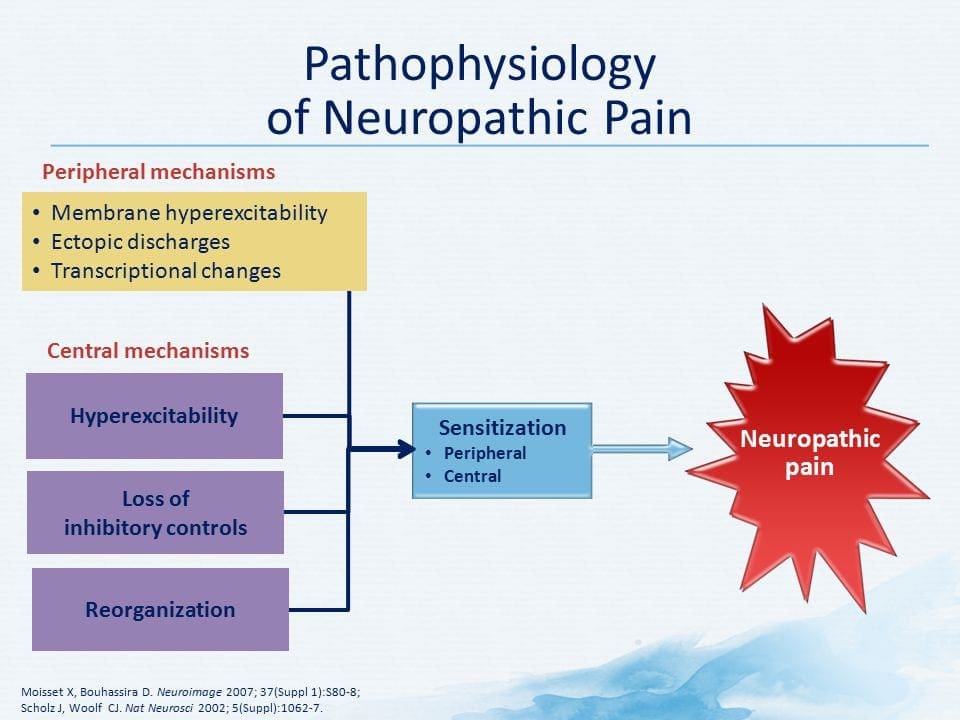
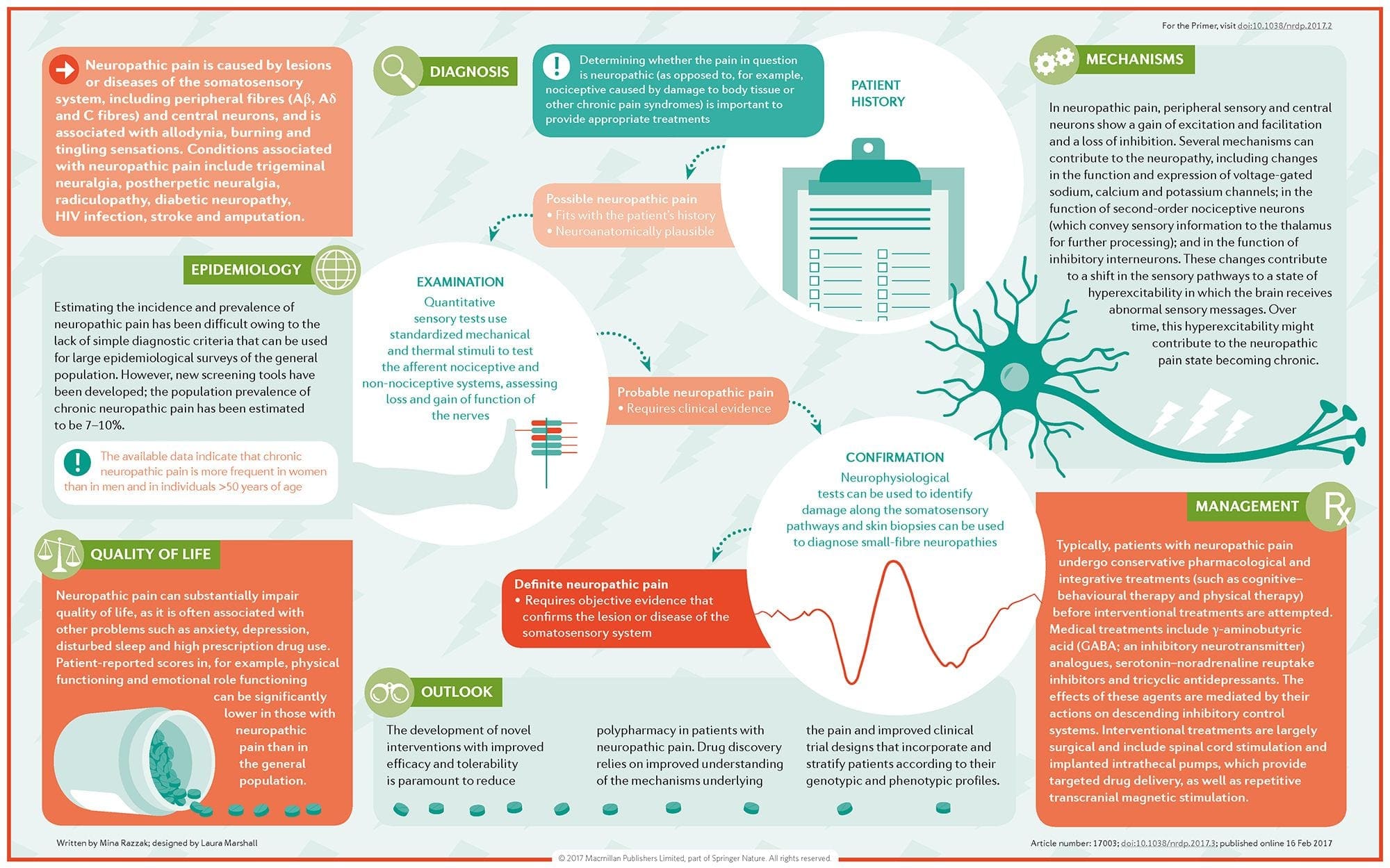



































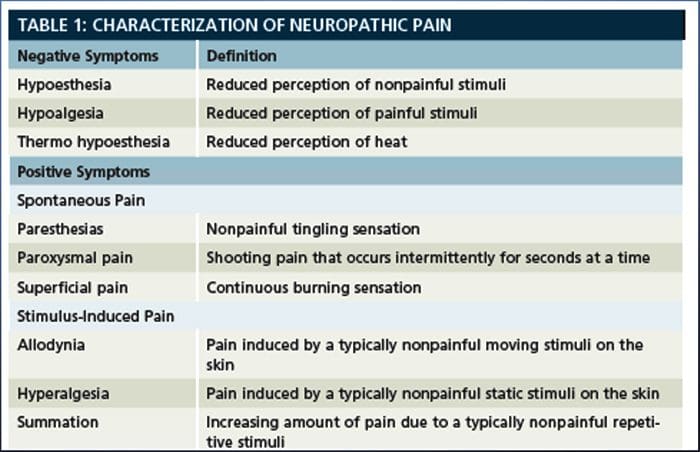
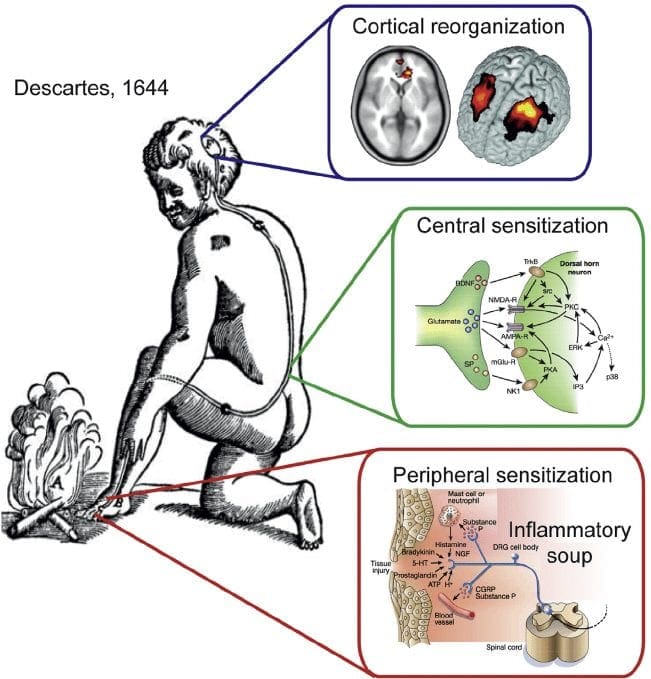
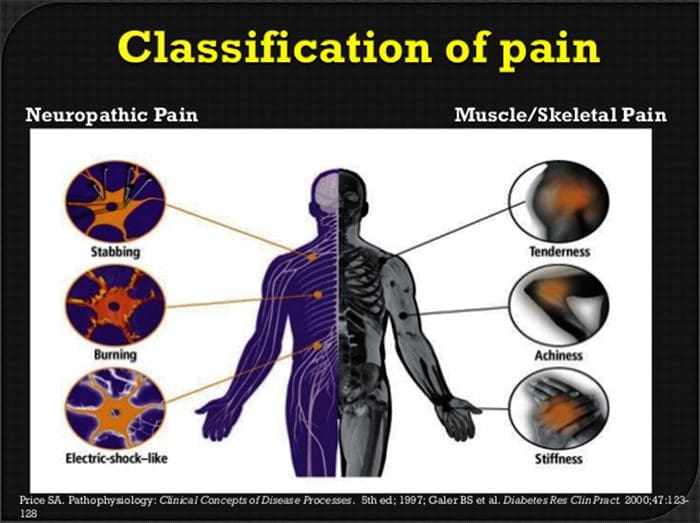
 ස්නායු රෝග වේදනාවේ ව්යාධිජනකය
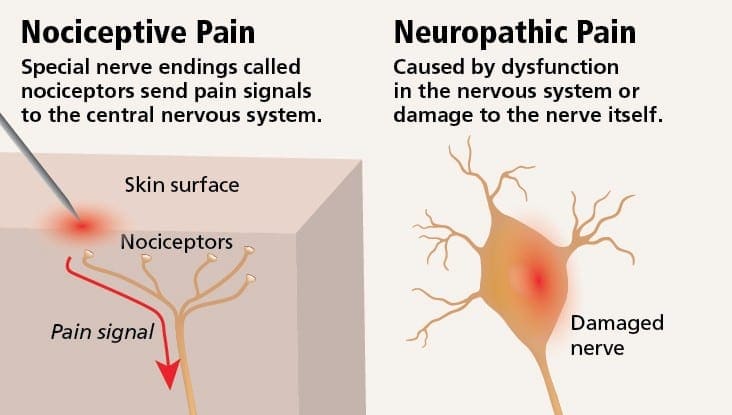
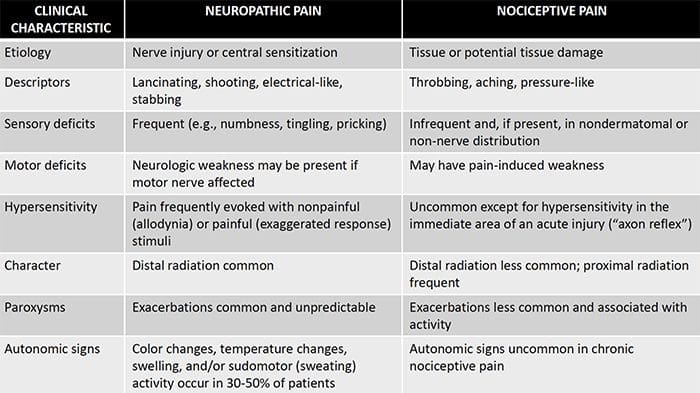
ස්නායු රෝග වේදනාවේ ව්යාධිජනකය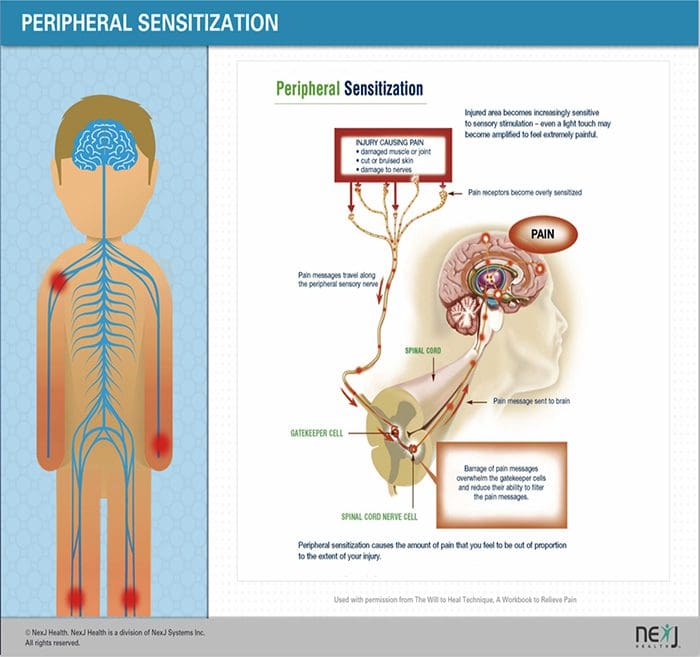
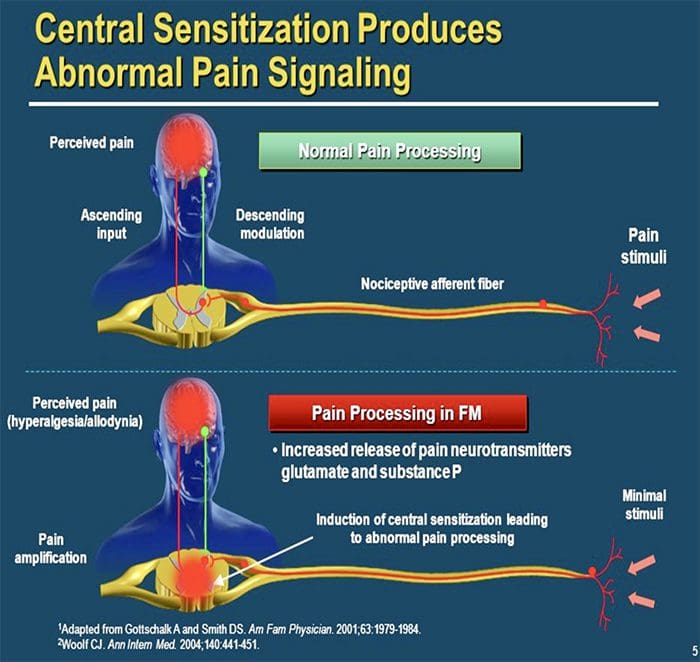
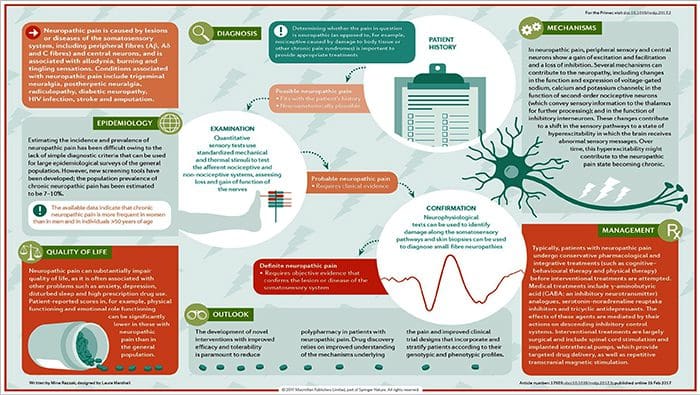
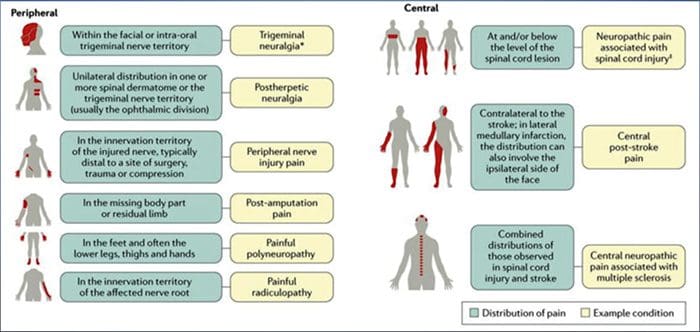 පොදු හේතු
පොදු හේතු





 PHANTOM LIMB PAIN & Augmented Reality
PHANTOM LIMB PAIN & Augmented Reality
 පොදු හේතු
පොදු හේතු



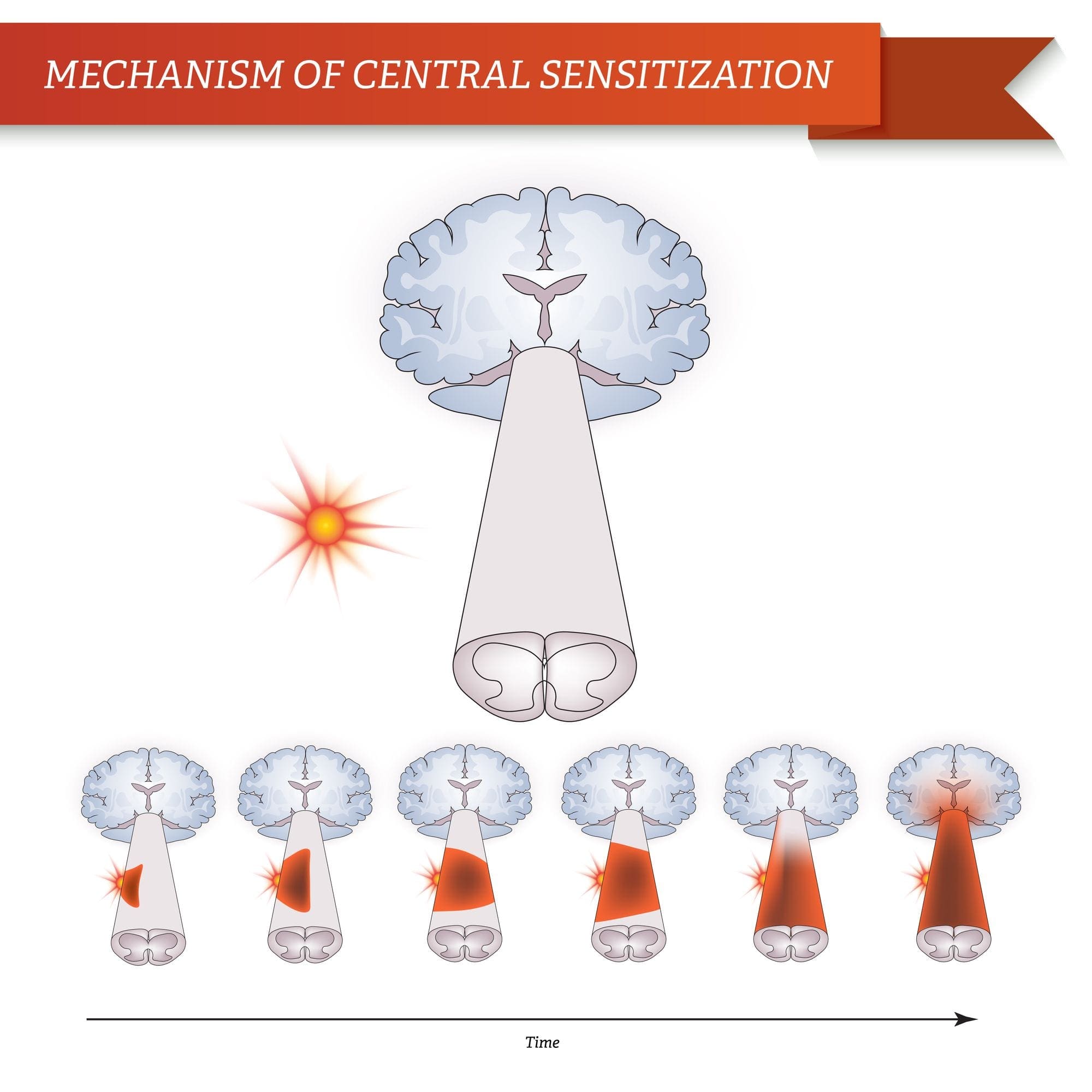
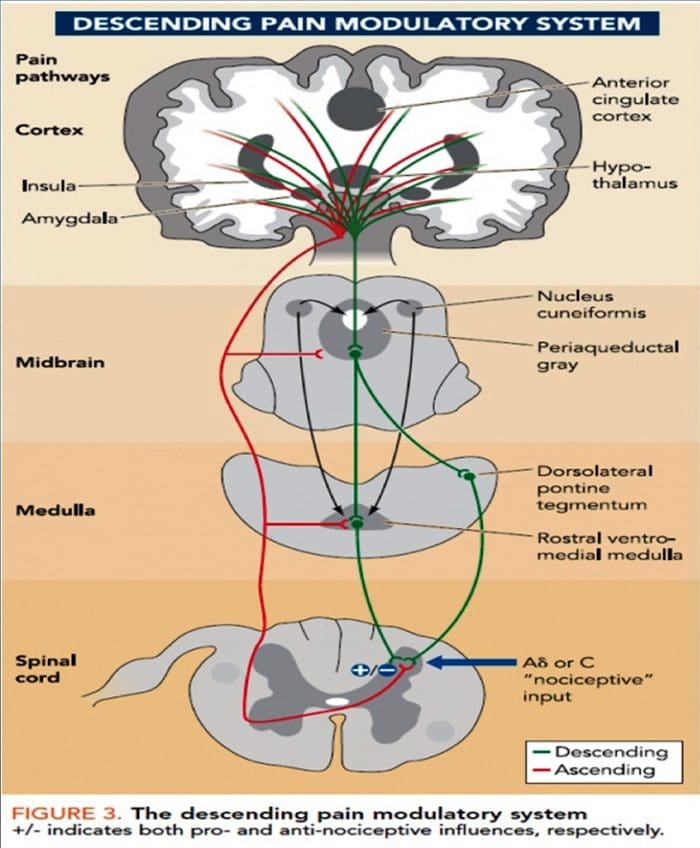
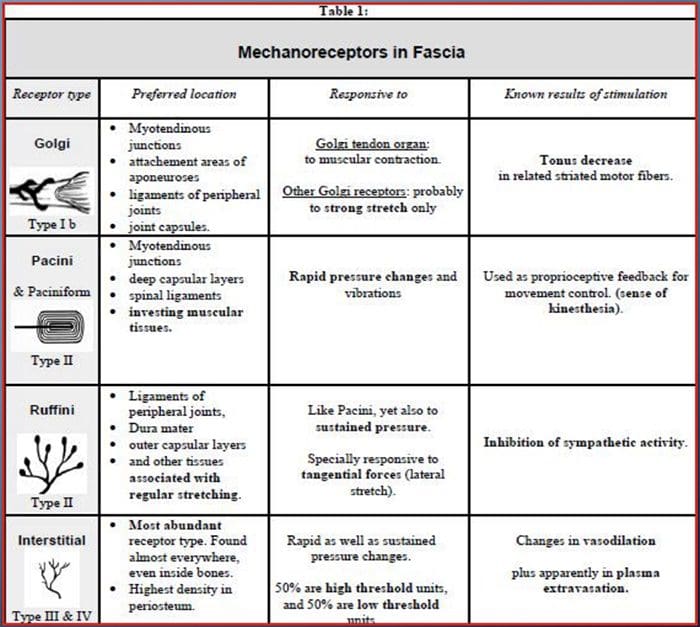 වේදනාව පිළිබඳ ගේට් පාලන න්යාය
වේදනාව පිළිබඳ ගේට් පාලන න්යාය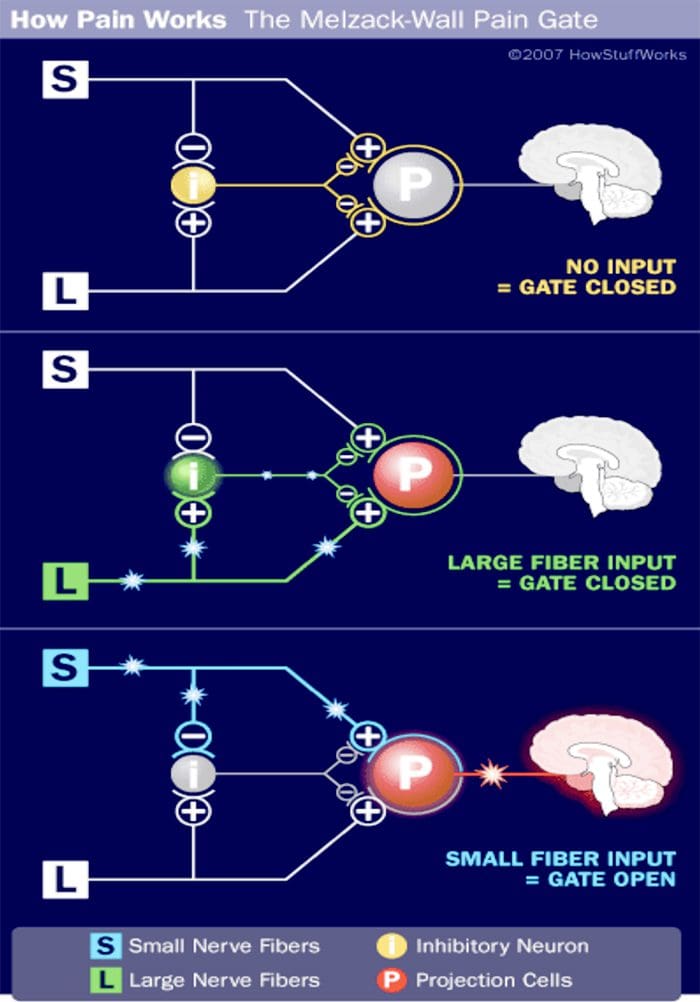
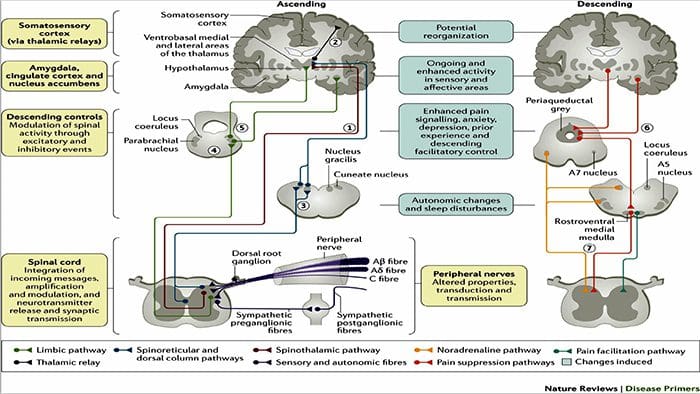
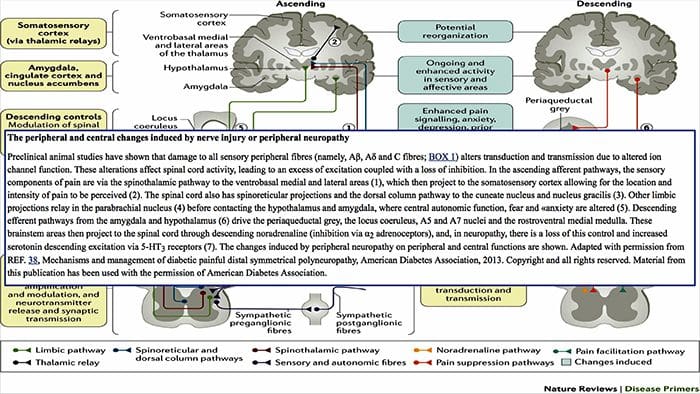
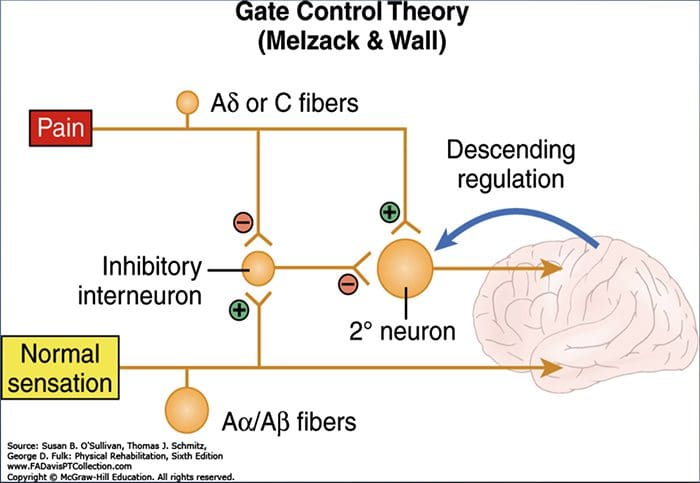 පෘෂ්ඨීය තීරු මධ්ය ලෙම්නිස්කල් මාර්ගය
පෘෂ්ඨීය තීරු මධ්ය ලෙම්නිස්කල් මාර්ගය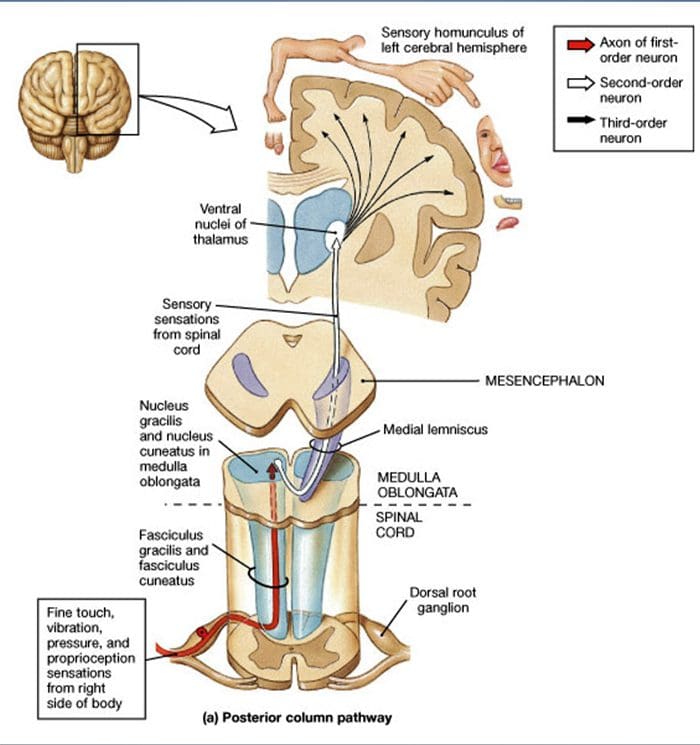 වේදනා සංජානනය වෙනස් කිරීම සඳහා පර්යන්ත යාන්ත්රික ග්රාහකයින් භාවිතා කිරීම
වේදනා සංජානනය වෙනස් කිරීම සඳහා පර්යන්ත යාන්ත්රික ග්රාහකයින් භාවිතා කිරීම
 විත්රීයකරණය
විත්රීයකරණය
 සැහැල්ලු ස්පර්ශය
සැහැල්ලු ස්පර්ශය



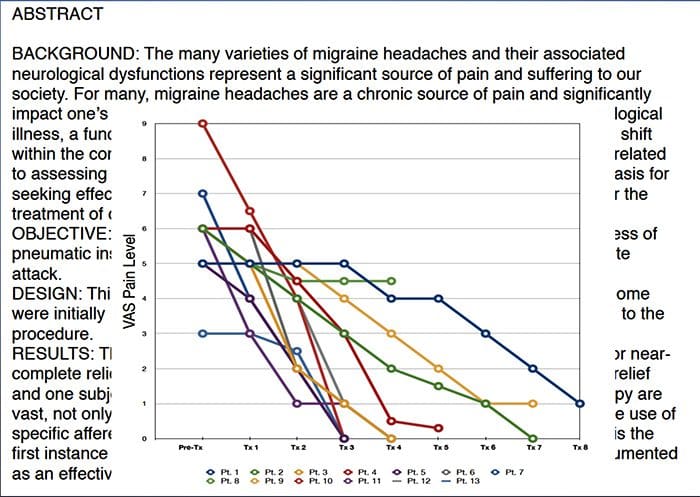

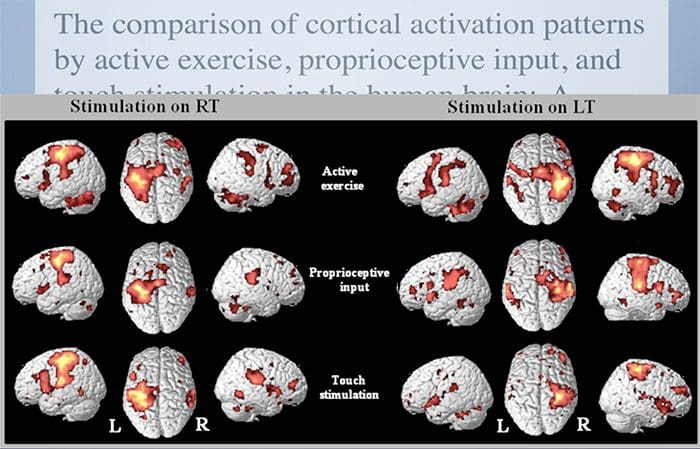
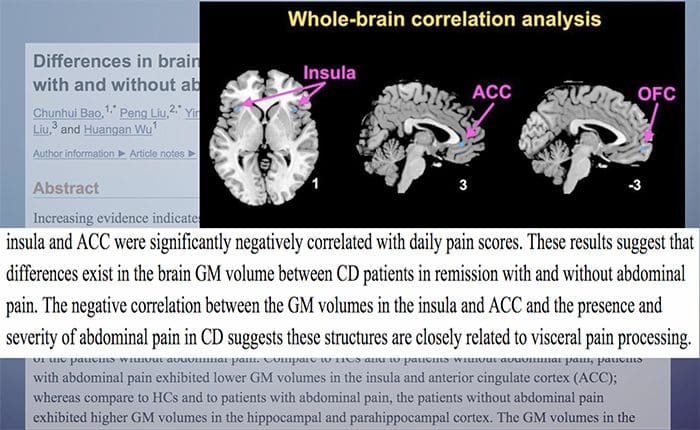

 CASE පාඩම්
CASE පාඩම්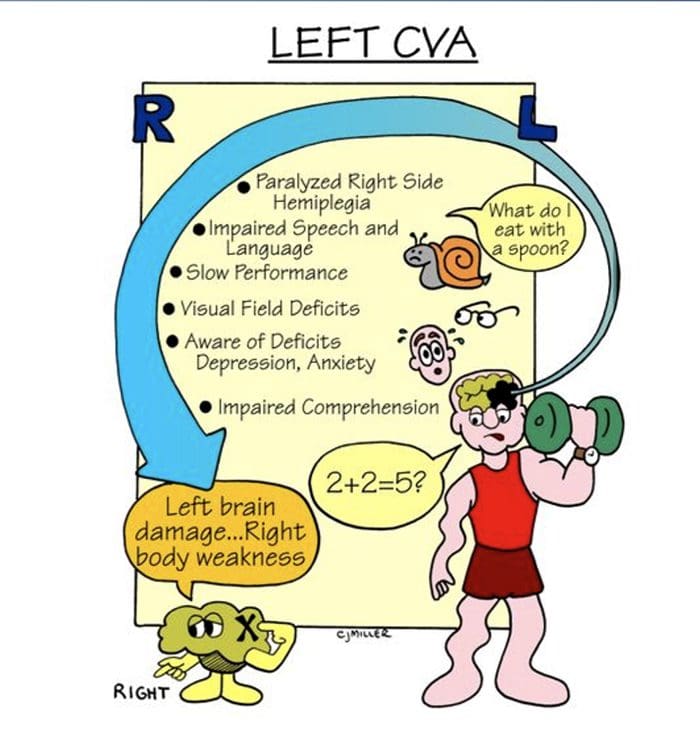 ශාරීරික විභාගයේ විශේෂ අවස්ථා
ශාරීරික විභාගයේ විශේෂ අවස්ථා ශාරීරික විභාගයේ විශේෂ අවස්ථා
ශාරීරික විභාගයේ විශේෂ අවස්ථා
 දින දෙකකට පසු
දින දෙකකට පසු